Heredity, Gene Regulation, and Development
I. Mendel's Contributions
II. Meiosis and
the Chromosomal Theory
III. Allelic, Genic,
and Environmental Interactions
IV. Sex Determination
and Sex Linkage
In the last lecture, we saw that
there were a variety of factors that influenced how an allele was expressed.
In those cases, however, the processes of Mendelian heredity - segregation and
independent assortment - still occurred as Mendel described. What gave us new
patterns was the way a genotype was expressed as a phenotype. In the next few
lectures, we will see that there are even deviations from the basic principles
of segregation and independent assortment.
A. Sex Determination
Although the sex of a human is typically
determined by the particular chromosomes received (XX for female and XY for
male), sex is determined in different ways in many other organisms. Before
we look at the consequences of chromosomally determined inheritance, let's put
it in perspective by looking at other ways that sex is determined. Here is a
fun article about a wide range of sexual
strategies.
1. Environmental Sex Determination
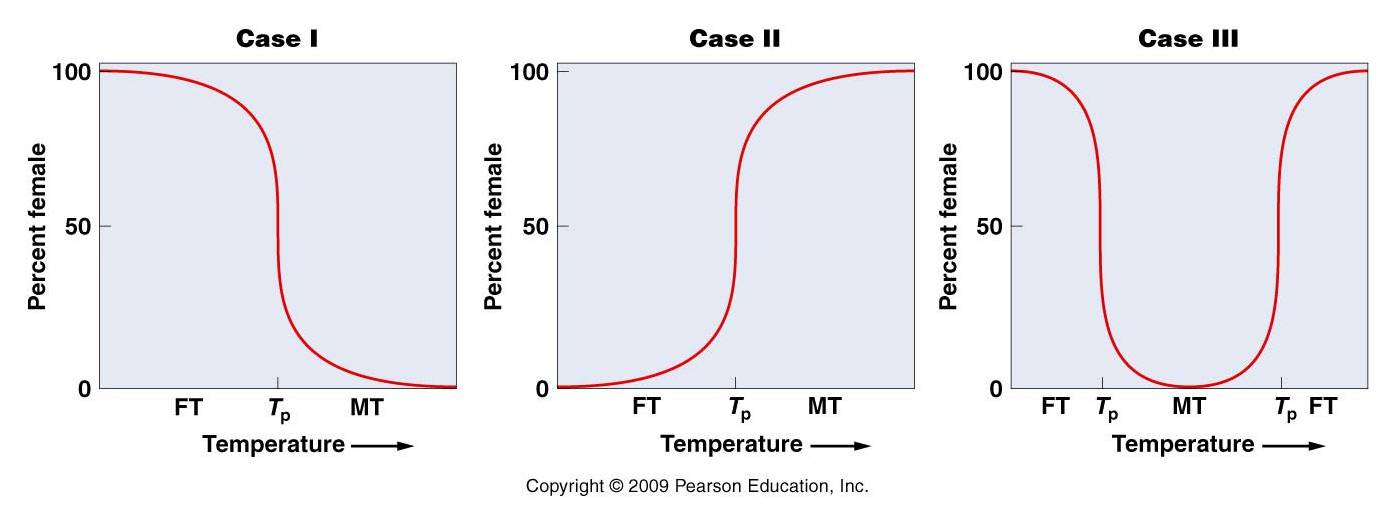 a.
temperature (in some
turtles, warm eggs develop into daughters and cool eggs into sons; in some lizards
and crocodilians, the pattern is reversed: cool = female, warm = sons). These
effects are not direct; what happens is that temperature differentially activates
sets of genes that influence certain hormonal pathways that affect the development
of gonads and genitalia. But these genes are not all on a given chromosome,
so there is no correlation between chromosomal complement or genotype and sex.
Species with temperature dependent sex determination don't change sex; sex is
determined once, by the temperature of incubation. Here
is a nice overview of the topic by Dr.
Michael Dorcas at Davidson.
a.
temperature (in some
turtles, warm eggs develop into daughters and cool eggs into sons; in some lizards
and crocodilians, the pattern is reversed: cool = female, warm = sons). These
effects are not direct; what happens is that temperature differentially activates
sets of genes that influence certain hormonal pathways that affect the development
of gonads and genitalia. But these genes are not all on a given chromosome,
so there is no correlation between chromosomal complement or genotype and sex.
Species with temperature dependent sex determination don't change sex; sex is
determined once, by the temperature of incubation. Here
is a nice overview of the topic by Dr.
Michael Dorcas at Davidson.
b. nutrients/size
In some species that
change sex, sex is determined by the size of the organism (and correlations
in the amount of resources they have available for reproduction). For example,
in some species like Jack-in-the-Pulpit (Arisaema triphyllum), small
plants are male and large plants are female. The adaptive benefits of this strategy
are easy to understand. It is cheap, energetically, to be a successfully reproducing
male in this species. Male plants just have to produce pollen (which produces
the sperm). It is more expensive, energetically, to be a female. Bigger gametes
are just the beginning; then you have to nurture the embryo and invest in additional
tissue, like fruit. So, if you are small, you can still be reproductively successful
as a male. But, you can't be as successful if you are small and female because
each offspring will cost you more energetically. So, selection has favored small
plants being male. So, why be female when you are big? Well the first reason
is a  probabilistic
argument.... if everyone else is male and you are the only female, then you
will mate and all of the offspring in the population will be yours (HUGE relative
reproductive success). However this reason is really unrelated to size. A second
reason is because you can adjust the QUALITY of your offspring. You can decide
to invest in all of them, or can invest in fewer and give them each more energy.
This give them a better chance of survival, which increases the likelyhood that
your genes (that are in these offspring), get passed on. So, if you are a female
plant and you have received pollen from many males, you can abort the embryos
that are doing poorly and only invest energy in embryos that are doing well.
This increases your reproductive success. And, this selective abortion doesn't
happen only in plants. European red deer (Cervus elaphus) have been
shown to selectively abort male embryos in years where resources are in short
supply. How can we understand this? Well, it is understandable in light of natural
selection and differential reproductive success in this harem-forming species.
Small male fawns grow into small male adults who won't mate - they won't acquire
a harem. Small does, however, still have a high probability of mating. So, consider
two starving, adult females that are pregnant with a male embryo and a felame
embryo, respectively. If they both bring their embryo to term and deliver it,
they will probably reduce the probability of their own survivorship (using the
last of their energy on their offspring) - so there is a cost. For the doe with
a male embryo, there is no fitness benefit; it's offspring won't mate. For the
doe with a female embryo, there will still be a reproductive payoff, as the
female offspring may still mate. Now, consider another starving doe with a male
embryo. If she aborts this embryo, she saves all the energy she would have spent
on growing that embryo to term. With thsi saved energy, she has a better chance
of surviving, herself - and reproducing, again. So, when resources are in short
supply, selection favors females that abort male embryos (and save the energy
to survive and breed again), but let female embryos develop (that are likely
to mate even though they are small). We
got a bit off track here - deer have chromosomally determined sex determination;
appreciate that this was just an example of selective abortion.
probabilistic
argument.... if everyone else is male and you are the only female, then you
will mate and all of the offspring in the population will be yours (HUGE relative
reproductive success). However this reason is really unrelated to size. A second
reason is because you can adjust the QUALITY of your offspring. You can decide
to invest in all of them, or can invest in fewer and give them each more energy.
This give them a better chance of survival, which increases the likelyhood that
your genes (that are in these offspring), get passed on. So, if you are a female
plant and you have received pollen from many males, you can abort the embryos
that are doing poorly and only invest energy in embryos that are doing well.
This increases your reproductive success. And, this selective abortion doesn't
happen only in plants. European red deer (Cervus elaphus) have been
shown to selectively abort male embryos in years where resources are in short
supply. How can we understand this? Well, it is understandable in light of natural
selection and differential reproductive success in this harem-forming species.
Small male fawns grow into small male adults who won't mate - they won't acquire
a harem. Small does, however, still have a high probability of mating. So, consider
two starving, adult females that are pregnant with a male embryo and a felame
embryo, respectively. If they both bring their embryo to term and deliver it,
they will probably reduce the probability of their own survivorship (using the
last of their energy on their offspring) - so there is a cost. For the doe with
a male embryo, there is no fitness benefit; it's offspring won't mate. For the
doe with a female embryo, there will still be a reproductive payoff, as the
female offspring may still mate. Now, consider another starving doe with a male
embryo. If she aborts this embryo, she saves all the energy she would have spent
on growing that embryo to term. With thsi saved energy, she has a better chance
of surviving, herself - and reproducing, again. So, when resources are in short
supply, selection favors females that abort male embryos (and save the energy
to survive and breed again), but let female embryos develop (that are likely
to mate even though they are small). We
got a bit off track here - deer have chromosomally determined sex determination;
appreciate that this was just an example of selective abortion.
 c.
Social Environment:
Many territorial reef fish like clown fish and wrasses have sequential hermaphrodism,
as we mentioned before. A dominant, large, mature individual (female clownfish,
male wrasse) dominates the other individuals in the school. In clownfish, all
but one of the other fish in the school remain juvenile males; the other developes
into a reproductive male who pairs with the dominant female. With the loss of
the mature dominant female, the mature male changes sex and becomes the dominant
female, one immature male developes into a mature male, and the others remain
sexually immature. In progynous wrasses, one female in a school developes into
a reproductive male who mates with the mature females. The loss of the male
stiumulates one of the females to change sex. The nature of these changes is
poorly understood, but in both cases the social dynamics in the school have
an effect. Aggressive behaviors by the dominant individual influence the hormone
concentrations in subordinate individuals, maintaining them as immatures or
in the other sex. Loss of this suppression (by the loss of the dominant individual)
releases one individual, who changes very quickly (often with associated morphological
and behavior changes) and assumes the aggressive, dominating behavior that continues
to suppress development in the other members of the school.
c.
Social Environment:
Many territorial reef fish like clown fish and wrasses have sequential hermaphrodism,
as we mentioned before. A dominant, large, mature individual (female clownfish,
male wrasse) dominates the other individuals in the school. In clownfish, all
but one of the other fish in the school remain juvenile males; the other developes
into a reproductive male who pairs with the dominant female. With the loss of
the mature dominant female, the mature male changes sex and becomes the dominant
female, one immature male developes into a mature male, and the others remain
sexually immature. In progynous wrasses, one female in a school developes into
a reproductive male who mates with the mature females. The loss of the male
stiumulates one of the females to change sex. The nature of these changes is
poorly understood, but in both cases the social dynamics in the school have
an effect. Aggressive behaviors by the dominant individual influence the hormone
concentrations in subordinate individuals, maintaining them as immatures or
in the other sex. Loss of this suppression (by the loss of the dominant individual)
releases one individual, who changes very quickly (often with associated morphological
and behavior changes) and assumes the aggressive, dominating behavior that continues
to suppress development in the other members of the school.
Relative size in a social setting
has direct effects on sex determination in the Midas cichlid (a fish). Within
a brood, larger individuals become male and smaller individuals become female
(this is permanent sex determination; there is no switching in these species).
By modifying the relative sizes of fish in a brood (adding larger fishes and/or
removing smaller fishes) the sexual 'fate' of the fish can be altered. So, it
is not size in an absolute sense (bigger than 5cm) that make a fish male, but
it is relative size - bigger than the other fish in the brood - that influences
male development (Francis and Barlow 1993).
This highlights an important point
about selection: SELECTION (AND EVOLUTION) DO NOT NECESSARILY ACT "FOR THE GOOD
OF THE SPECIES". Selection maximizes the immediate reproductive success among
individuals. Curiously, what's good for an individual might be bad for the long-term
success of the population as a whole. For instance, having only 1 female in
the population REALLY inhibits the growth rate of the population. The population
will be alot smaller than if you had many females all producing offspring. And,
smaller populations are closer to extinction, so any random event might bounce
the population size to zero. But, selection FAVORS females that can inhibit
the female development of their peers. (Like it favors male deer that can inhibit
the mating of their male peers). So it happens, even if it is "bad" for the
species in the long run. If an individual (male OR female) can usurp all of
the mating (or a disproportionate share of the mating) in a population for themselves,
then they will reproduce more than other members of the population and these
genes for usurping control will increase in frequency. So, in harem forming
species, males that keep other males from mating have the most offspring, and
the male offspring inherit a belligerent attitude that stimulates then to fight
other males and try to keep them from mating. You see, it is not necessarily
the TOTAL number of offspring that selection favors. It is the number of offspring
produced RELATIVE TO other members of the population that selection favors.
So, if a male can keep other males from mating, even if he only has 1 offspring,
he will still be doing better than the other males in the population and will
be at a selective advantage (his genes will increase in frequency relative to
theirs). Likewise, if a female can inhibit the development of other females,
then all of the offspring produced in the population - even if fathered by different
males - will necessarily be hers and the genes for inhibiting the development
or reproduction of other females will be passed on in the population.
2. Chromosomal
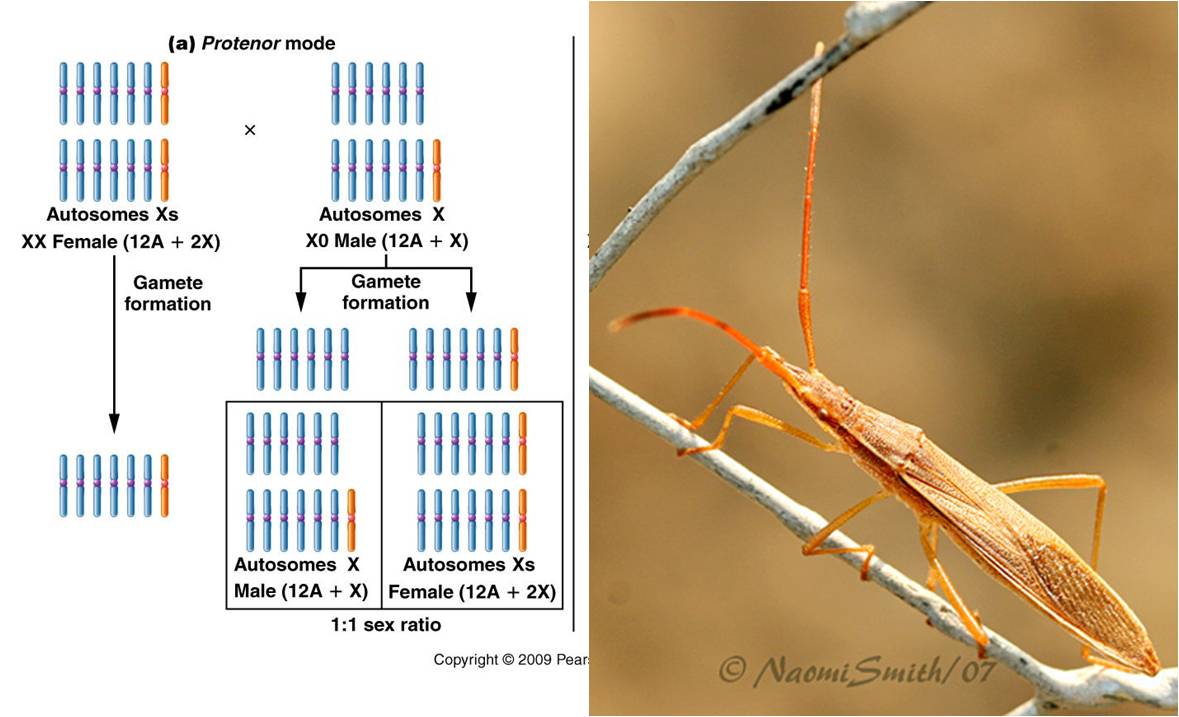 a.
Protenor (a
Homopteran... a true bug) - A single sex chromosome - 6 autosomal, no sex chromosome
(female)
a.
Protenor (a
Homopteran... a true bug) - A single sex chromosome - 6 autosomal, no sex chromosome
(female)
6 autosomes, plus sex chromosome (male)
b.
Lygaeus (another bug) - two sex chromosomes:
- In humans and most mammals, the heterogametic sex is male (XY) and the homogametic
sex if female (XX). In humans and most mammals, the presence of a "y" chromosome
causes male development, even if the individual has 2 X's by mistake (XXY).
At least, usually individuals that are XY are male. Actually, 'maleness'
is determined by only a small region of the Y - called the "sex determining
region of the y" - SRY - gene. This single gene can be lost from the chromosome
through mutation, and sometimes it can even be transferred to the X during male
sperm production. So, this man produces "y" sperm that LACK the sry, and "X"
sperm that have the sry. (This only occurs in the couple sperm where this mutation
occurred; it would not occur in all this man's sperm. So, this is VERY rare.
This sry "X" chromosome, when paired with the X of the mother, would produce
a zygote that is XX but has the sry and would develop into a male child. Likewise,
if a sperm with the mutant Y is used, then a zygote is produced that is XY but
has no sry gene and developes into a female.
- In birds, some reptiles, and butterflies
and moths, the homogametic sex is male (WW) whereas the heterogametic sex (ZW)
is female. We use the Z W symbols to represent sexes in these species.
c. Balanced
sex determination
In Drosophila fruit flies,
it is the ratio of X chromosomes to autosomal sets that determines the sex of
the organism. If the ratio is 1:1, like 2 X chromosomes in a normal diploid
organism with two sets of autosomes, then the organism developes as a female.
If the ratio is 1:2, with only a single X chromosome and 2 sets of autosomes,
the organism is a male. This seems very much like Lygaeus determination,
and for normal diploid XX (female) and XY (male) flies, it is. But the difference
was demonstrated by identifying the sexes of mutants that lack or gain an extra
sex chromosome. So, for instance, humans that are 45, X (2 sets of autosomes
but one only sex chromosome, so the total number of chromosomes is 45 instead
of 46) are female - because they lack the Y. In flies, however, diploid organisms
with only one X develop as males, even though they lack a Y. In another example,
a human that is 47, XXY (with an extra X and thus, 47 total chromosomes) would
be male because of the presence of a Y. However, a fly that is diploid and has
XXY sex chromosomes will be female (ratio of autosomal sets: X's = 1:1).
B. Gender
 Gender
is a role or behavior that a society correlates with a sex. These 'gender-roles'
and behaviors vary across societies, even though most humans are biologically
male or female. So, in the USA, we might associate the wearing of make-up and
dressing in skirts as playing the gender role of a woman. In the Wodaabe tribe
of the Sahel region of Africa, these cultural practices are associated with
the gender-role of a man. And don't dare tell a scotsman he is wearing a skirt...
it's a kilt! In some societies, the expected 'social norm' of sexual preference
for someone playing the gender role of a man is to be attracted to women. In
ancient Greece, sexual behavior was not as strongly correlated with our gender
categories of 'man' and 'woman', but by the categories of the 'active role'
(adult, penetrative, maculine, higher social status) and 'passive role' (youth,
feminine, lower social status). These roles could be played by members of the
same or opposite sex.
Gender
is a role or behavior that a society correlates with a sex. These 'gender-roles'
and behaviors vary across societies, even though most humans are biologically
male or female. So, in the USA, we might associate the wearing of make-up and
dressing in skirts as playing the gender role of a woman. In the Wodaabe tribe
of the Sahel region of Africa, these cultural practices are associated with
the gender-role of a man. And don't dare tell a scotsman he is wearing a skirt...
it's a kilt! In some societies, the expected 'social norm' of sexual preference
for someone playing the gender role of a man is to be attracted to women. In
ancient Greece, sexual behavior was not as strongly correlated with our gender
categories of 'man' and 'woman', but by the categories of the 'active role'
(adult, penetrative, maculine, higher social status) and 'passive role' (youth,
feminine, lower social status). These roles could be played by members of the
same or opposite sex.
 In
almost all social animals, there are correlations between sex and social behavior.
But, as in humans, these behaviors can vary between populations and between
closely related species. Bonobo chimps, for instance, mate face-to-face like
humans, and are different from common chimps to whom they are more closely related.
Indeed, sex is used as a form of communication in bonobos - it has become a
ritualized behavior of communication within and between sexes, often used as
a greeting between any two bonobos, male or female. Many other species also
engage in frequent sexual behaviors between members of the same sex; 90% of
observed sexual pairings in giraffes occurs between males. So, sex in giraffes
may be like sex in Ancient Athens - a way to communicate dominance. Or, as in
bonobo's, it may just be done for fun and social bonding. There are lots of
cases where organisms use sex for reasons other than pro-creation. And, in almost
all social species, there is a fraction of individuals that choose same-sex
pairings, or perform the behavioral gender roles typical for the other sex.
In some species, individuals even use gender roles to deceive others in their
own population. For example, in cuttlefish (relatives of squid) young males
color themselves as females and enter the territory of dominant males. They
play the gender role of a female, and are allowed into the territory. When the
dominant male isn't looking, the immature male serruptitiously mates with females
in the territory. These males are called 'sneaker' males. The same thing happens
in many birds, where young males maintain their immature, female-like plumage
and gain access to male territories where they mate with females. As with nearly
everything we've examine in biology, a more rigorous examination of patterns
in nature tend to reveal continua; even between categories that, at the outset,
seem discrete: categories like species, sexes, gender, and sexual behavior.
In
almost all social animals, there are correlations between sex and social behavior.
But, as in humans, these behaviors can vary between populations and between
closely related species. Bonobo chimps, for instance, mate face-to-face like
humans, and are different from common chimps to whom they are more closely related.
Indeed, sex is used as a form of communication in bonobos - it has become a
ritualized behavior of communication within and between sexes, often used as
a greeting between any two bonobos, male or female. Many other species also
engage in frequent sexual behaviors between members of the same sex; 90% of
observed sexual pairings in giraffes occurs between males. So, sex in giraffes
may be like sex in Ancient Athens - a way to communicate dominance. Or, as in
bonobo's, it may just be done for fun and social bonding. There are lots of
cases where organisms use sex for reasons other than pro-creation. And, in almost
all social species, there is a fraction of individuals that choose same-sex
pairings, or perform the behavioral gender roles typical for the other sex.
In some species, individuals even use gender roles to deceive others in their
own population. For example, in cuttlefish (relatives of squid) young males
color themselves as females and enter the territory of dominant males. They
play the gender role of a female, and are allowed into the territory. When the
dominant male isn't looking, the immature male serruptitiously mates with females
in the territory. These males are called 'sneaker' males. The same thing happens
in many birds, where young males maintain their immature, female-like plumage
and gain access to male territories where they mate with females. As with nearly
everything we've examine in biology, a more rigorous examination of patterns
in nature tend to reveal continua; even between categories that, at the outset,
seem discrete: categories like species, sexes, gender, and sexual behavior.
C. Sex Linkage
Sex linkage describes the pattern
of heredity between traits governed by genes on the sex chromosomes and the
trait of 'sex', itself. In humans, although the Y is a small chromosome with
little besides the sry gene, the X is a large chromosome with hundreds of genes
that govern a wide variety of phenotypic traits. Since the X chromosome influences
sex, and also carries genes that influence these traits, the pattern of heredity
of sex and these X-linked genes is not independent; whereever the X goes (and
influences sex), these other genes go, too. So,
the pattern of heredity of these genes is NOT independent of sex - violating
the pattern of independent assortment between sex and these x-linked traits.
1. For Comparision:
The Independent Assortment of Sex (as a trait) and an Autosomal Trait:
Genes that are not on sex chromosomes
are called autosomal genes. So, suppose we followed the pattern of heredity
of "tongue rolling" and sex in humans. Tongue rolling is the ability
to curl your tongue into a tube, by lifting the edges of your tongue up. It
is a trait that is dominant ('A') to the inability to roll the tongue ('a').
Because this trait is autosomal, it is governed by genes on a pair of homologous
chromosomes that are NOT the sex chromsomes. So, the way the autosomes pair
during gamete formation will be independent of how the sex chromosomes pair,
and the genes they carry will be inherited independently of one another. So,
suppose we do reciprocal dihybrid crosses:
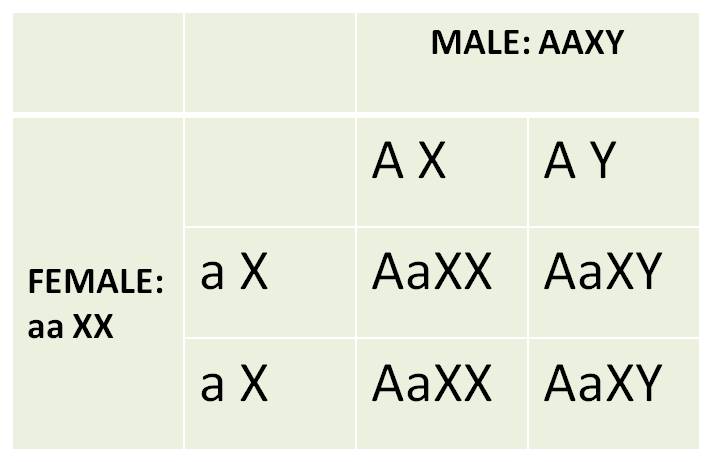 |
 |
100%
tongue rolling in sons and daughters |
100%
tongue rolling in sons and daughters |
As expected, the results of the reciprocal
crosses are the same, and are consistent with Mendel's predictions for independently
assorting traits; the sex of the tongue-rolling parent doesn't matter - results
in the reciprocal crosses are the same. Now, let's see what happens when we
consider two traits governed by genes on the same chromosomes - in this case,
with sex-linked genes, one trait is "sex" and the other trait is colorblindness.
2. An Example
of Sex-Linkage: Red-Green Colorblindness in Humans:
There are many forms of 'color blindness'.
Two of these are caused by alleles on the X chromosome. In one case, the red-detecting
photoreceptors are absent, and a person sees red as black. In another form,
the green photoreceptors are absent, affect the ability to distinguish red from
green. As with all X linked traits, these conditions are expressed in males
more frequently than in females. When males inherit an X with these alleles,
they must express the trait; they have no other chromosome that carries these
genes, because as males, their other sex chromsomes is a Y that does not carry
these genes. So, consider this set of reciprocal crosses:
 |
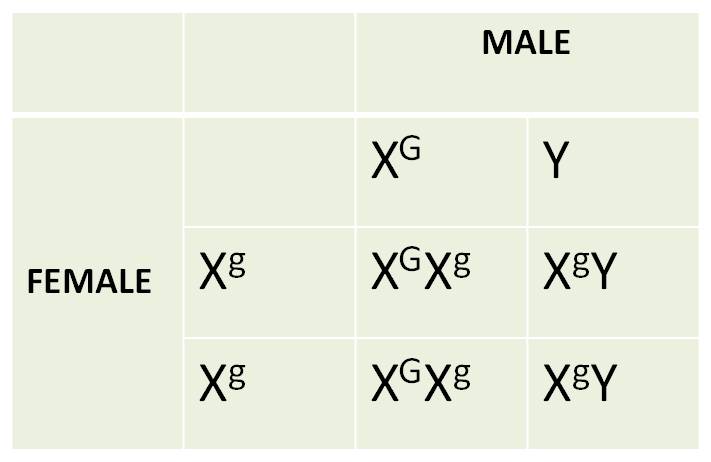 |
100%
normal vision |
50%
colorblind (all males): 50% normal (all females) |
The results of the reciprocal cross
are DIFFERENT. Unlike Mendel's experiments where the sex of the parent giving
a trait didn't matter, here it does; sons look like their mothers. Even when
the mother does not express the trait, but carries a recessive allele in the
heterozygous condition, there is a 50% chance that her son will inherit the
allele on their X and expressing the trait. Males with normal vision will only
have daughters with normal vision, because they will pass their one normal X
to all their daughters.
Study Questions
1. Explain why females are often larger than
males, even in species in which individuals change sex.
2. Provide an example of social sex determinism.
3. Describe Protenor and Lygaeus sex determination.
4. What are 'homogametic' and 'heterogametic'
sexes?
5. What is 'gender'?
6. How do animals use sex, beside procreation?
7. Describe a circumstance where members of one
sex in animals accept the 'gender role' of the other sex.
8. Explain why sex linkage violates the priciple
of independent assortment.
2. Consider this cross: AaXBY
x AaXBXb
- How many types of gametes can the male
make with respect to these loci?
- How many phenotypes are possible in the
progeny (Assume IA and complete dominance for both loci).
- What fraction of male offspring will
express the Ab phenotype?