
Mutation is defined as a change in the genome. Mutations can occur due to damage to the DNA by mutagens or by errors in DNA replication. If these changes are not repaired before division, they are passed to daughter cells. Mutations occuring in body cells are called 'somatic mutations', and (unless they occur in cells that will become ovaries or testes) the effects will not be passed to offspring. However, mutations can also occur in reproductive tissues - in the cells that will produce gametes (or spores), or in the process of meiosis, itself. These changes can be inherited by offspring, and are called 'germline mutations'. Obviously, if one of these mutant gametes is invovled in a fertilization event, then the mutation is present in the zygote and in ALL CELLS of the resulting offspring. These heritable changes are our focus.
The changes can occur at four broad levels: 1) changing the number of chromosomal sets (polyploidy); 2) changing the number of chromosomes in a set (aneuploidy); 3) changing the number and arrangement of genes on chromosomes (duplications, deletions, translocations); and 4) changing the nitrogenous base sequence of a gene. ALL of these changes are mutations, and we will look at each level of changes in order - from large changes in chromosome number to small changes in nitrogenous base sequence.
For the most part, mutations occur randomly. And as you would suspect, making random changes to a complex, functional system like a living cell are probably going to have a negative effect. This is particularly true for large changes, like the gain or loss of individual chromosomes or chromosome sets. In humans and other animals with complex development and tissue specialization, most of the large-scale changes cause such profound problems with development that the embryo does not develop correctly and is spontaneously aborted. Over 90% of all spontaneous abortuses (that are tested) have chromosomal anomalies. By analogy, think about the complex system of an automobile. If you make a large, random change to a functioning car - like taking out the engine, or putting one big wheel on the front right axle - it is unlikely that you will improve the functioning of the system. Small changes are probably going to have a negative effect, also, but it is more likely that a small change has a beneficial effect than a large change. For example, you might randomly change the angle of the rear-view mirror every time you get in the car. Well, sometimes you will throw it off so you can't see, but sometimes, just by chance, you will make a small change that adjusts it to a better viewing angle - even if it was already pretty good.
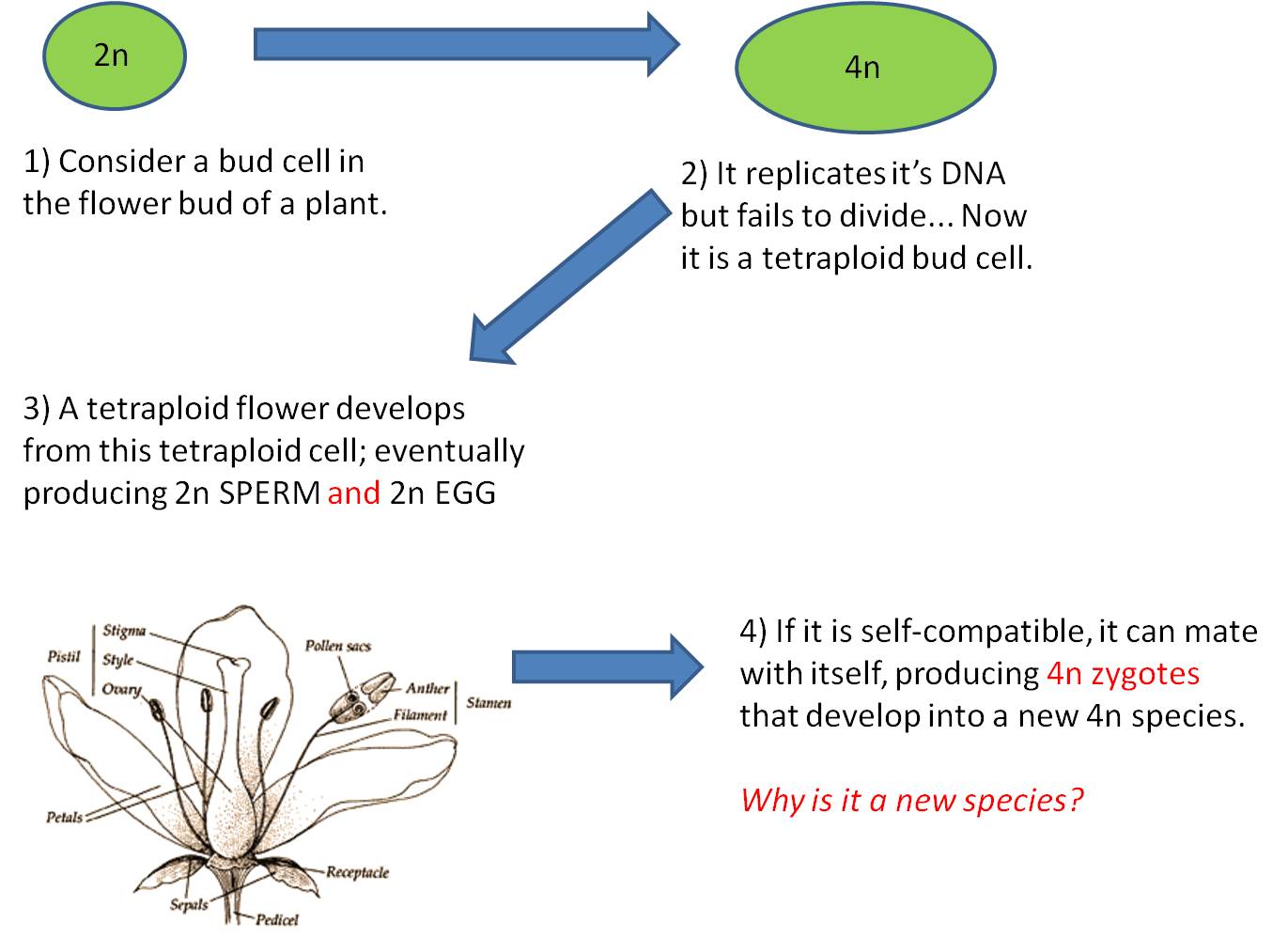
This is the most dramatic change possible; an addition of an entire SET of chromosomes, changing a haploid gamete to a diploid gamete, or changing a diploid cell into a tetraploid cell.
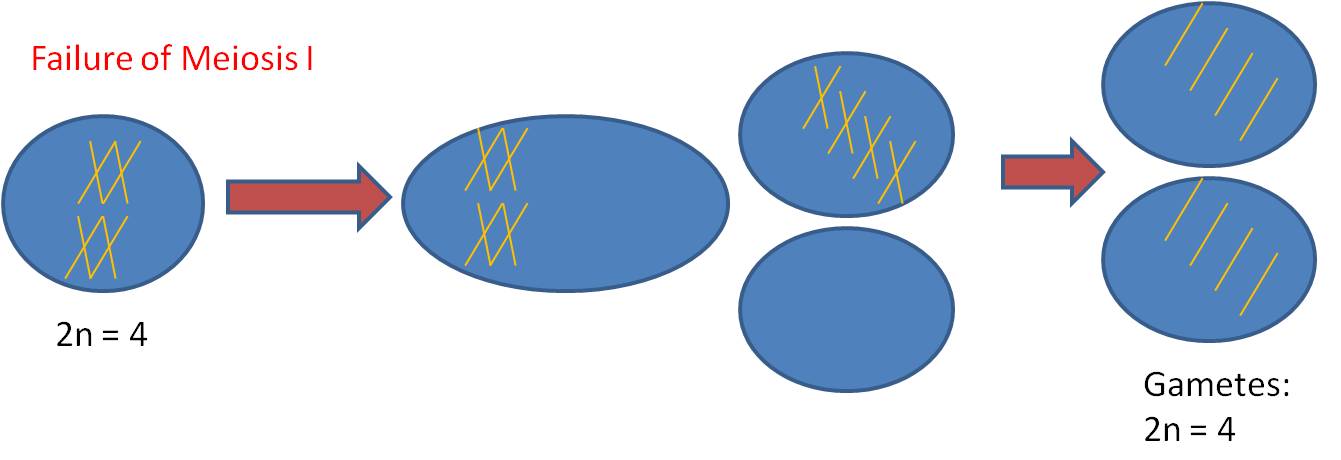 1.
Mechanism #1 - Complete failure of Meiosis
1.
Mechanism #1 - Complete failure of Meiosis
a. process
b. result
 2.
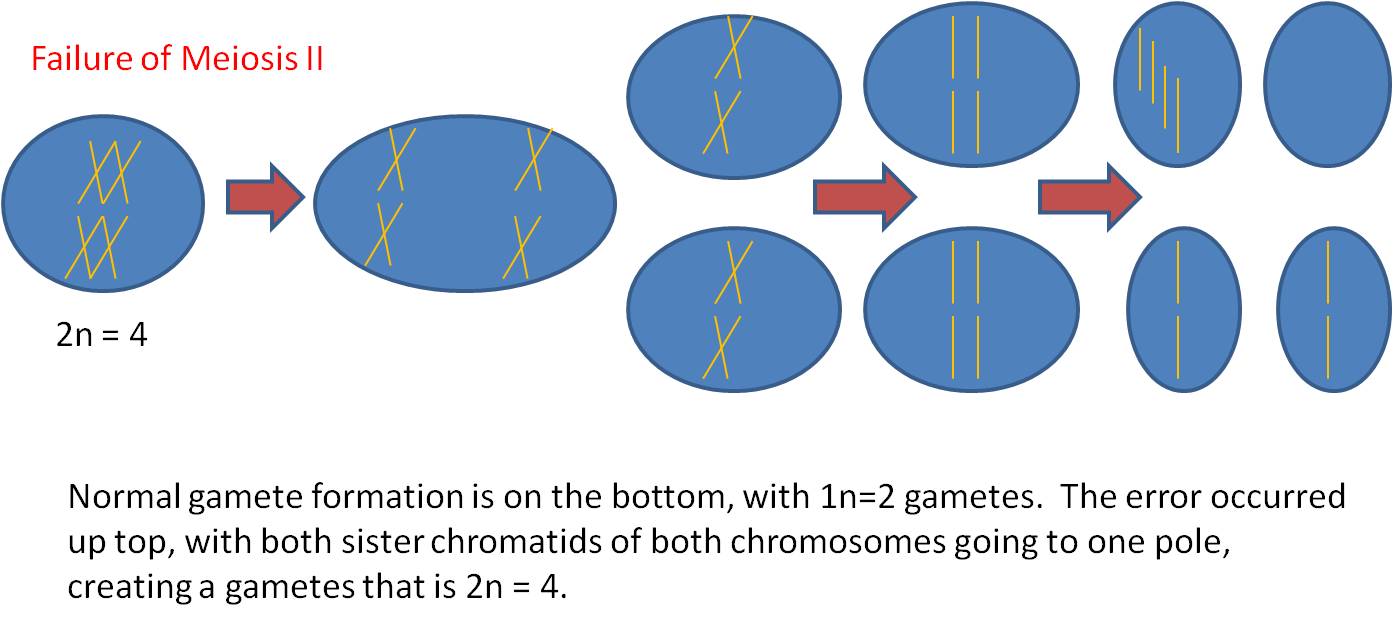
Mechanism #2 - Failure of Mitosis
2.
Mechanism #2 - Failure of Mitosis
Here, however, there are 2n male AND 2n female gametes produced. If the plant is self-fertile, these sperm can fertilize these eggs and a 4n zygote will be produced. This is TETRAPLOIDY. - Tetraploid offspring may not survive; there may still be enough of a disruption in the protein concentrations to screw-up development. But, if they can survive, then they WILL be able to make gametes by normal meiosis. Their 4n cells CAN divide evenly, producing 2n gametes.
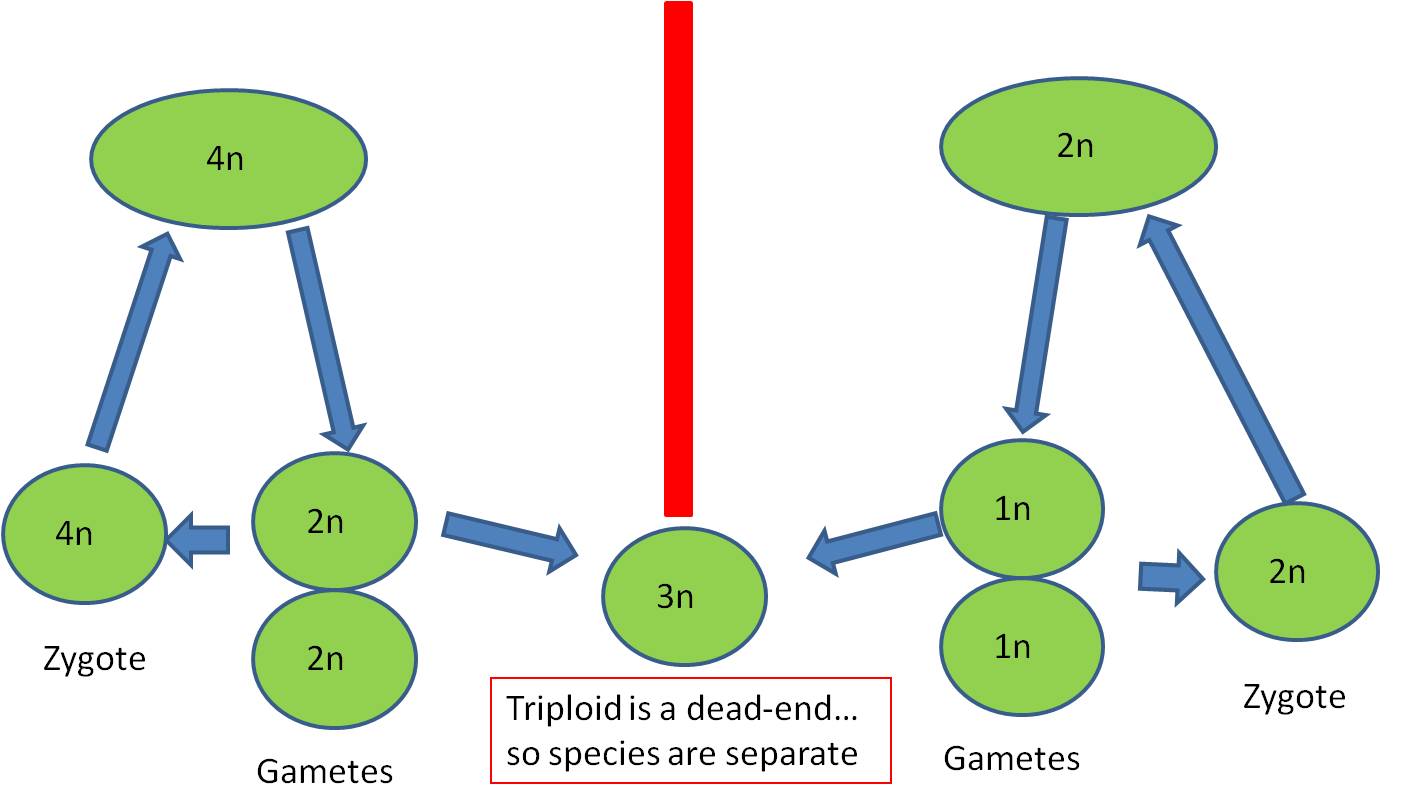
- This lineage could reproduce itself, and would be reproductively isolated from its originating species because IF their 2n gametes fertilized the haploid gametes of the originating species, a 3n triploid would be produced, which is typically a reproductive dead-end for reasons described, above.
 3.
Frequency of Polyploidy
3.
Frequency of Polyploidy
Polyploidy (>2n) is VERY rare in birds and mammals. The only known polyploid mammal is a small tetraploid mouse that lives in the Andes - it was discovered in 2002. No birds are known to be triploids. However, it is more frequent (but still rare) in fish, amphibians, and reptiles, where there are parthenogenetic 3n sister species of normal 2n species. However, on the whole, polyploidy is rare in dioecious species (where individuals are EITHER male or female). This is probably because the most likely polyploid product in these species is a triploid, which usually has the sterility problems mentioned earlier.
In hermaphroditic species (and particularly self-compatible plants), polyploidy is FAR more common. This is probably because tetraploidy is far more likely, and the tetraploid will be fertile. In fact, about 30-50% of all flowering plant species are polyploid!!!. That means that this mechanism has been a pretty important source of new species. Some genera have species with sequential increases in ploidy. For instance three different species of goldenrods have 14, 28, and 56 chromosomes, respectively. So, a reasonable hypothesis is that the second and third species were formed through polyploid events (failure in mitosis) as described above - doubling their ploidy levels and making new species.
 This
refers to the loss or addition of a single chromosome, not whole sets of chromosomes.
This mutation is called aneuploidy. 'Ploidy' refers
to set, 'eu' means true, and 'an' means not. So,
you have "not a true set".
This
refers to the loss or addition of a single chromosome, not whole sets of chromosomes.
This mutation is called aneuploidy. 'Ploidy' refers
to set, 'eu' means true, and 'an' means not. So,
you have "not a true set".
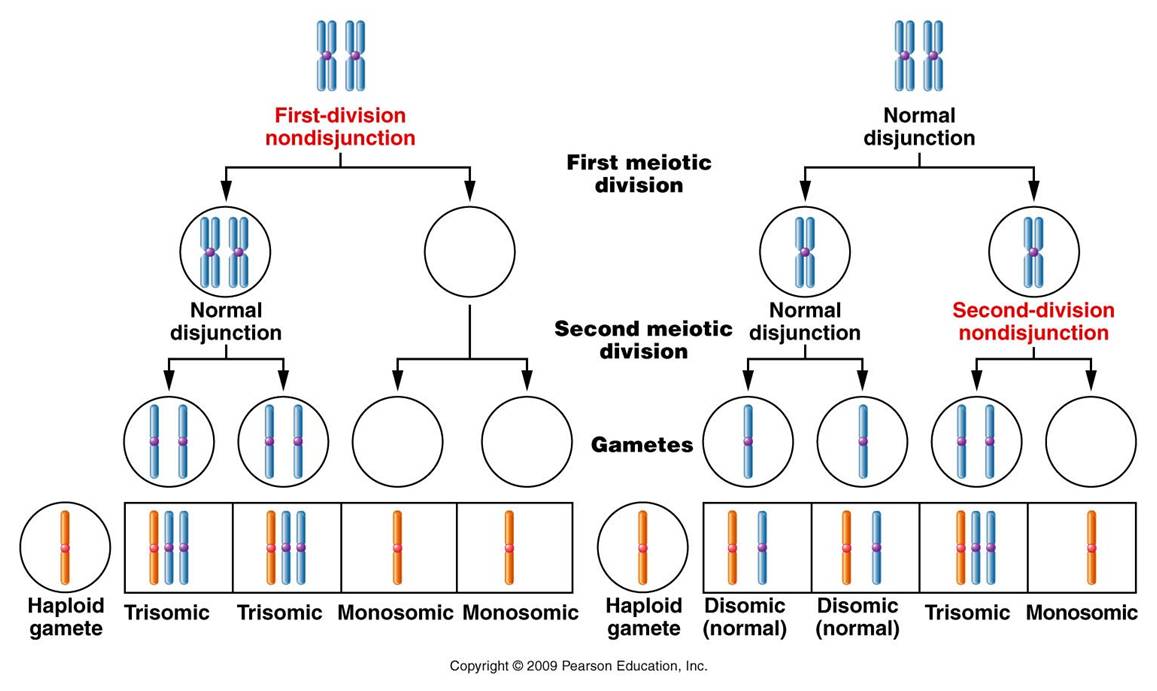
1. Mechanism:
Non-disjunction - During meiosis, a pair of homologous chromosomes (or sister chromatids) fail to segregate; both go to one daughter cell. So, we end up with a gamete with an extra chromosome, and a gamete that is one chromosome short.
When fertilization occurs with a normal haploid gamete, we end up with a zygote with an extra chromosome - a third chromosome in one homologous 'pair'. (Remember, one gamete inherited BOTH homologs, and then the other parent correctly contributes a single chromosome for this set.) So, this situation is called trisomy (three bodies.) If the other aberrant gamete is involved in a fertilization event, we end up with a zygote with only one chromosome in a given homologous set - this is a monosomy.
2. Human Conditions:
 In humans, these
imbalances are almost always fatal to the developing embryo - they are usually
spontaneously aborted during development (>90% of spontaneous abortions have
chromosomal anomalies). However, some embryos with these conditions can
complete development to birth, and some individuals can survive for decades.
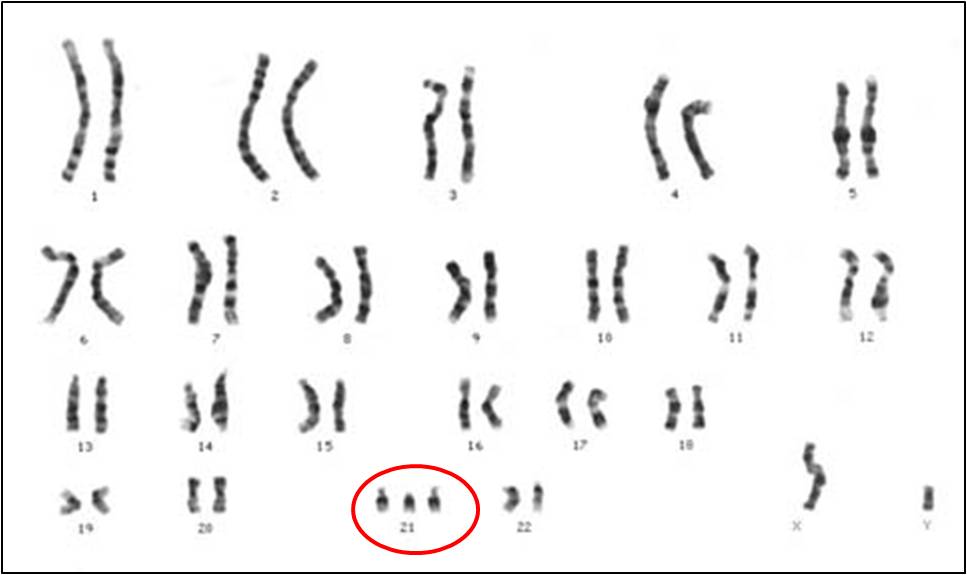
When non-disjunction occurs in the #21 chromosome, TRISOMY 21 occurs. This is
also known as Down's Syndrome. Other autosomal trisomies can survive to birth,
but that is rare and the effects are much more severe. Trisomy 13, Patau Syndrome,
can result in cyclopian eye development (among a host of other deadly anomalies).
Edward's Syndrome, trisomy 18, also causes a rash of typically lethal deformities.
Trisomies in the sex chromosomes are tolerated more often, resulting in XXX,
XXY, and XYY offspring. It is important to appreciate that Down's and sex trisomies
are the trisomies with the LEAST severe effects, where the individual survives
birth and lives for decades. Trisomies should, by chance, be just as likely
for other homologous sets, too; but apparently these trisomies are so devastating
to development that they result in spontaneous abortion.
In humans, these
imbalances are almost always fatal to the developing embryo - they are usually
spontaneously aborted during development (>90% of spontaneous abortions have
chromosomal anomalies). However, some embryos with these conditions can
complete development to birth, and some individuals can survive for decades.
When non-disjunction occurs in the #21 chromosome, TRISOMY 21 occurs. This is
also known as Down's Syndrome. Other autosomal trisomies can survive to birth,
but that is rare and the effects are much more severe. Trisomy 13, Patau Syndrome,
can result in cyclopian eye development (among a host of other deadly anomalies).
Edward's Syndrome, trisomy 18, also causes a rash of typically lethal deformities.
Trisomies in the sex chromosomes are tolerated more often, resulting in XXX,
XXY, and XYY offspring. It is important to appreciate that Down's and sex trisomies
are the trisomies with the LEAST severe effects, where the individual survives
birth and lives for decades. Trisomies should, by chance, be just as likely
for other homologous sets, too; but apparently these trisomies are so devastating
to development that they result in spontaneous abortion.
In humans, the only human monosomy that does, periodically, survive to birth and beyond is the monosomy for the sex chromosome, with 1 X chromosome. This condition, 45, XO, is called Turner's syndrome. No other monosomies are tolerated in humans; all are spontaneously arborted.
1. Gene Duplicatin and Deletion:
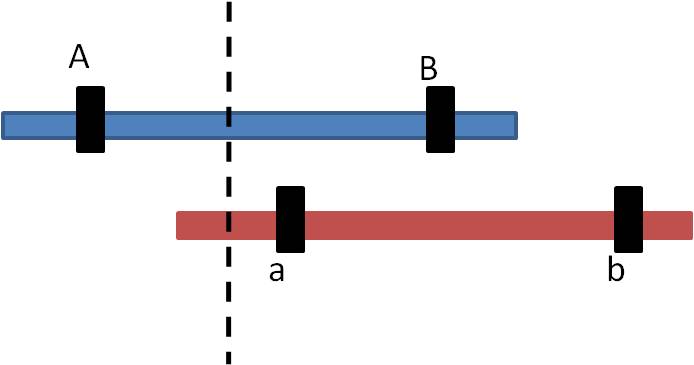
a. process-unequal crossing-over:
Sometimes during Prophase I of meiosis, the homologs do not line up evenly. If a cross-over event occurs as marked, then the pieces of DNA that are exchanged are not equivalent.
 So, the products look like this:
So, the products look like this: 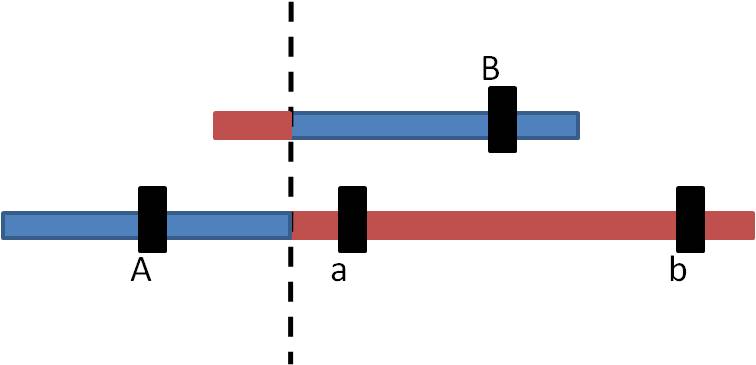
- The "A" locus has been DUPLICATED on the bottom chromosome, and DELETED from the top chromosome.
b. effects:
- Can be bad: DELETION of genes is usually bad...
the gene probably did something useful, and now it is gone. At the least,
organisms that inherit a deletion MUST express the gene at that lacus that they
received from the other parent. So, deletions result in the increased
frequency of expression of deleterious recessives inherited from the other parent.
That's bad, too. (This is similar to why males expressed X-linked recessives
more frequently than females - because they only get one X chromosome, so whatever
is on it gets expressed. In that case, the pattern is true for ALL genes on
the X chromosome. Here, with a deletion (that can occur on any chromosome),
the effect is limited to the genes on the deletion region. But, whatever the
organism inherits from the other parent must be expressed.)
DUPLICATION can also be bad. By having three active genes in an other wise
diploid genome, the concentration of proteins produced by the duplicated gene
may now be "out of whack" (a technical term) with the concentrations
of other proteins. Essentially, it is similar to the effects of a trisomy, but
at a smaller genetic scale. - Can be good: Duplication of a gene can be good
for two reasons. First, maybe more is better. As we mentioned before,
there are many genes for r-RNA production, and many genes for melanin production.
How did we get so many genes that are all the same, and code for the same thing?
It would be VERY unlikely for random mutation, occurring in two different places
in the genome, to create the same sequence of nitrogenous bases just by chance.
So, how did it happen? Well, you just saw how - unequal crossing over. And if
'more is better', then organisms with more copies of a gene will be selected
for, and these genomes will increase in frequency in the population. 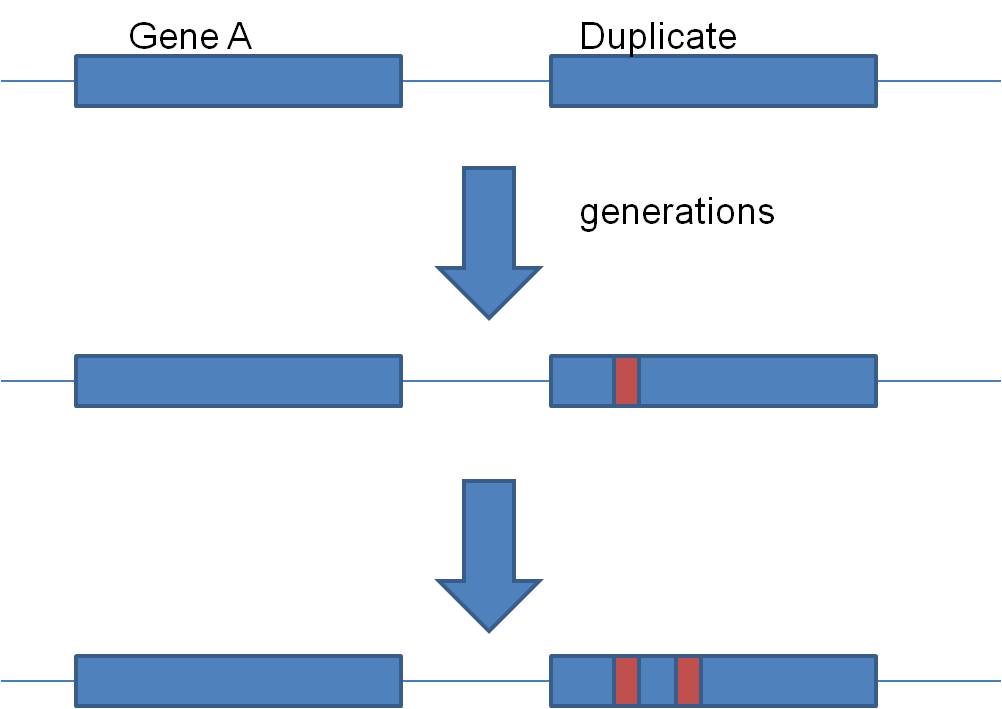 Second, gene duplication is an important source of EVOLUTIONARY NOVELTY - new,
useful variation. You see, it is easier to get something NEW and FUNCTIONAL
by modifying something that already works, rather than building it from scratch.
So, by duplicating a functional gene, evolution can now introduce mutations
into that "copy" and make something new. By tweaking that gene, we might
get something new that also works, but in a slightly different way. This would
produce a "family" of genes that all produce the same type of protein, but proteins
that work a bit differently. We call these families "gene families", and
they are quite common. A classic example is the "globin family" of genes.
This includes the alpha, beta, gamma, and myo- globin genes. They are
all similar enough to suggest common ancestry from one ancestral gene, yet different
enough to do slightly different things. And they are all similar enough
in nucleotide sequence that their origin by duplication and subsequent slight
modification is strongly indicated.
There is ANOTHER reason why duplication is an important
source of evolutionary novelty, and it relates to an "intelligent design" criticism
of evolution. Proponents of intelligent design and creationism suggest
that evolution by gradual change is impossible if the intermediate steps are
non-functional. For instance, how could the beta-globin gene "evolve from"
an alpha globin gene by a gradual process if the intermediate steps are non
functional? You see, here's the problem:
Second, gene duplication is an important source of EVOLUTIONARY NOVELTY - new,
useful variation. You see, it is easier to get something NEW and FUNCTIONAL
by modifying something that already works, rather than building it from scratch.
So, by duplicating a functional gene, evolution can now introduce mutations
into that "copy" and make something new. By tweaking that gene, we might
get something new that also works, but in a slightly different way. This would
produce a "family" of genes that all produce the same type of protein, but proteins
that work a bit differently. We call these families "gene families", and
they are quite common. A classic example is the "globin family" of genes.
This includes the alpha, beta, gamma, and myo- globin genes. They are
all similar enough to suggest common ancestry from one ancestral gene, yet different
enough to do slightly different things. And they are all similar enough
in nucleotide sequence that their origin by duplication and subsequent slight
modification is strongly indicated.
There is ANOTHER reason why duplication is an important
source of evolutionary novelty, and it relates to an "intelligent design" criticism
of evolution. Proponents of intelligent design and creationism suggest
that evolution by gradual change is impossible if the intermediate steps are
non-functional. For instance, how could the beta-globin gene "evolve from"
an alpha globin gene by a gradual process if the intermediate steps are non
functional? You see, here's the problem: 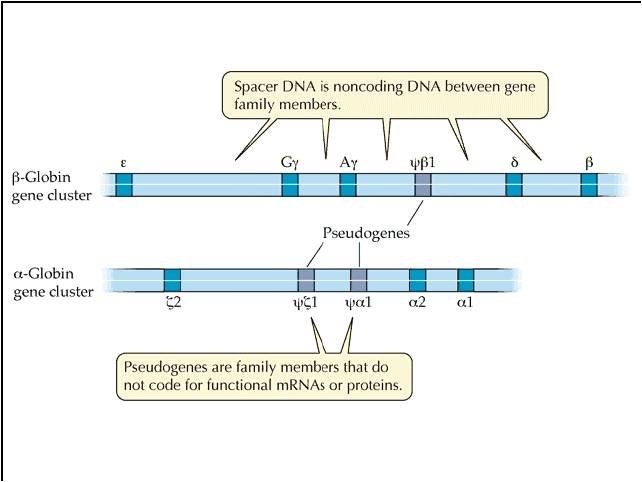 Suppose alpha
globin differs from beta globin by 5 amino acids (I have no idea how many -
this is just an example). In order for the beta globin gene to "evolve",
5 different mutations in the DNA would have to occur, in order to code for five
new amino acids. Now, it is VERY unlikely that all five of those particular
mutations would occur at the same time - mutations are very rare. BUT
(argue the creationists), that's exactly what would have to happen for beta
globin to evolve from alpha globin. Otherwise, they argue, ff we have
one mutation at a time, the alpha globin is turned off and we have NO alpha
globin and NO beta globin - a non-functional intermediate that should be selected
against.
BUT, duplication solves this apparent dilemma. If the alpha gene duplicates,
there are now TWO alpha globin genes. Suppose the first mutation occurs
in one, and shuts that gene off. So what? The other gene is still
right there, cranking out the functinal alpha globin protein. So, the organism
with the non-functional duplication is NOT selected against. Rather, they pass
their chromosome on to offspring, with the functional and non-functional genes.
Generations later, the 2nd, 3rd, 4th, and 5th mutations can accumulate.
When the 5th happens, the product is functional beta globin.... and now the
organisms that have inherited this mutation have both functional alpha and functional
beta globin. Gene duplication is a VERY important mechanism of introducing
evolutionarily SIGNIFICANT variation, because it maintains the original gene
and its function, while allowing mutations to "play" with the other gene.
Mutations that turn that genes off are not selected against, but mutations that
improve the gene/protein can still have a positive effect and be selected for.
In fact, many regions of non-coding DNA in our genome are "pseudogenes"....
regions of non-functional DNA that bear a remarkable similarity in nucleotide
sequence to a functional gene. The hypothesis is that they represent a
duplicated gene that has since been turned off by mutation.
Suppose alpha
globin differs from beta globin by 5 amino acids (I have no idea how many -
this is just an example). In order for the beta globin gene to "evolve",
5 different mutations in the DNA would have to occur, in order to code for five
new amino acids. Now, it is VERY unlikely that all five of those particular
mutations would occur at the same time - mutations are very rare. BUT
(argue the creationists), that's exactly what would have to happen for beta
globin to evolve from alpha globin. Otherwise, they argue, ff we have
one mutation at a time, the alpha globin is turned off and we have NO alpha
globin and NO beta globin - a non-functional intermediate that should be selected
against.
BUT, duplication solves this apparent dilemma. If the alpha gene duplicates,
there are now TWO alpha globin genes. Suppose the first mutation occurs
in one, and shuts that gene off. So what? The other gene is still
right there, cranking out the functinal alpha globin protein. So, the organism
with the non-functional duplication is NOT selected against. Rather, they pass
their chromosome on to offspring, with the functional and non-functional genes.
Generations later, the 2nd, 3rd, 4th, and 5th mutations can accumulate.
When the 5th happens, the product is functional beta globin.... and now the
organisms that have inherited this mutation have both functional alpha and functional
beta globin. Gene duplication is a VERY important mechanism of introducing
evolutionarily SIGNIFICANT variation, because it maintains the original gene
and its function, while allowing mutations to "play" with the other gene.
Mutations that turn that genes off are not selected against, but mutations that
improve the gene/protein can still have a positive effect and be selected for.
In fact, many regions of non-coding DNA in our genome are "pseudogenes"....
regions of non-functional DNA that bear a remarkable similarity in nucleotide
sequence to a functional gene. The hypothesis is that they represent a
duplicated gene that has since been turned off by mutation.
There is also another bit of evidence here. We often see members of gene families
in the same region of a chromosome - just as we would expect from unequal crossing-over
that places a duplicate next to its homologous gene. So, both the sequence similiarity
and the spatial proximity that we often see in gene families is strong evidence
that unequal cross-over and gene duplication have been important sources of
new useful genes.
1. Mechanism #1: Crossing Over within eukaryotic
genes can produce new combinations of exons, and new alleles: - In eukaryotes (and some prokaryotes) with the exon-intron structure
of their genes, crossing over WITHIN A GENE, in non-coding intron regions, can
create new combinations of exons. This may be a particularly productive
way of creating new functional alleles. Why? Well, consider two
alleles, A and a, that both function. For simplicity, suppose they are
composed of 3 exons each: 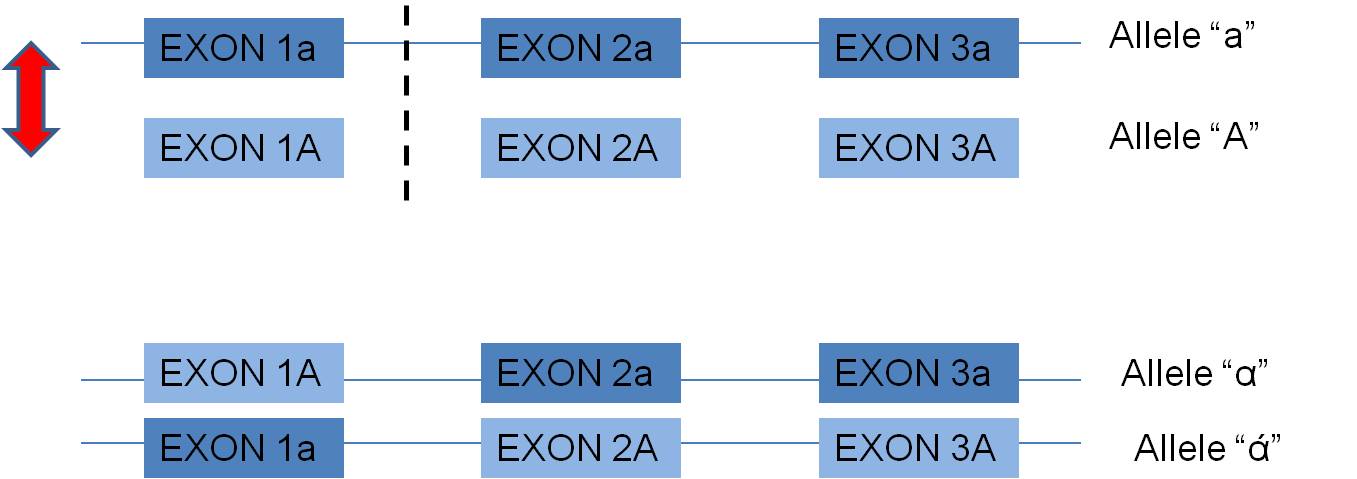 Now, the exons already function with one another. A cross-over between
the first and second exon may produce new combinations of exons that also have
a high probability of functioning, but maybe in a slightly new way. In
fact, many genes across the genome seem to share particular exons that encode
for a particular functional domain of proteins. It is far more likely
that these are homologous similiaries arising from "exon shuffling" than the
independent but parallel creating of these identical sequences by mutation.
Now, the exons already function with one another. A cross-over between
the first and second exon may produce new combinations of exons that also have
a high probability of functioning, but maybe in a slightly new way. In
fact, many genes across the genome seem to share particular exons that encode
for a particular functional domain of proteins. It is far more likely
that these are homologous similiaries arising from "exon shuffling" than the
independent but parallel creating of these identical sequences by mutation.
2. Mechanism #2: Point Mutations - changes in nucleotide
sequence a.
Addition/deletion: When DNA
is being replicated, the DNA polymerase may erroneously "skip" a base, or erroneously
add an extra base into the newly forming strand. If this is the sense
strand that will later be transcribed, this will cause the addition or deletion
of a base in the m-RNA. This change will "shift" the 3-base reading frame
during translation, changing every codon downstream:  So, from the point of the addition or deletion, every amino acid that follow
will change. So, these "frameshift" mutations usually have a deleterious
effect because they change a whole bunch of amino acids. b.
Substitutions: - during DNA replication, an improper base can be substituted while the new strand is being synthesized. Many of these errors are corrected
during 'DNA proof-reading' and repair that occurs in G1 and G2, but some (1
in a million) slip by. This change in the DNA (if in an exon) would change one
base in the m-RNA, and change one m-RNA codon. This could change an amino acid - but if you look at the genetic code you will see that
most AA's are encoded by more than one three base codon. So, for instance,
CUU, CUA, CUG, and CUC all code for the amino acid "leucine". So,
a change in the third position of the DNA sequence encoding these codons would
have NO EFFECT on the amino acid:
Original DNA = GAT ....... original RNA = CUA ....... amino acid = leucine
So, from the point of the addition or deletion, every amino acid that follow
will change. So, these "frameshift" mutations usually have a deleterious
effect because they change a whole bunch of amino acids. b.
Substitutions: - during DNA replication, an improper base can be substituted while the new strand is being synthesized. Many of these errors are corrected
during 'DNA proof-reading' and repair that occurs in G1 and G2, but some (1
in a million) slip by. This change in the DNA (if in an exon) would change one
base in the m-RNA, and change one m-RNA codon. This could change an amino acid - but if you look at the genetic code you will see that
most AA's are encoded by more than one three base codon. So, for instance,
CUU, CUA, CUG, and CUC all code for the amino acid "leucine". So,
a change in the third position of the DNA sequence encoding these codons would
have NO EFFECT on the amino acid:
Original DNA = GAT ....... original RNA = CUA ....... amino acid = leucine
mutant DNA = GAA ........ mutant RNA = CUU ...... amino
acid is STILL LEUCINE
So, some mutations are "silent", even if they occur within an exon of a gene.
However, other mutations can change the amino acid:
Original DNA = GAT ....... original RNA = CUA ....... amino acid = leucine
mutant DNA = GTT ........ mutant RNA = CAA ...... amino
acid is GLYCINE  The smallest change that is possible in DNA is a single nucleotide, and this
may cause the smallest change possible in a protein - a change in a single amino
acid. These small changes may be deleterious, neutral, or beneficial.
And, AS YOU KNOW, the value of a gene may depend on the environment.
Consider the sickle cell anemia example that we discussed before. Hemoglobin
consists of two alpha globin molecules and two beta globin molecules.
The allele for the normal beta globin chain contains 146 amino acids, with glutamine
as the 6th amino acid. The sickle cell hemoglobin contains the SAME alpha
chains as normal hemoglobin, but a mutant form of the beta chain. In the
mutant form, the 6th amino acid is valine. This difference in one amino acid
is caused by a change in one DNA nucleotide. As we have discussed, it
is beneficial in the tropics but deleterious in the temperate zone.
The smallest change that is possible in DNA is a single nucleotide, and this
may cause the smallest change possible in a protein - a change in a single amino
acid. These small changes may be deleterious, neutral, or beneficial.
And, AS YOU KNOW, the value of a gene may depend on the environment.
Consider the sickle cell anemia example that we discussed before. Hemoglobin
consists of two alpha globin molecules and two beta globin molecules.
The allele for the normal beta globin chain contains 146 amino acids, with glutamine
as the 6th amino acid. The sickle cell hemoglobin contains the SAME alpha
chains as normal hemoglobin, but a mutant form of the beta chain. In the
mutant form, the 6th amino acid is valine. This difference in one amino acid
is caused by a change in one DNA nucleotide. As we have discussed, it
is beneficial in the tropics but deleterious in the temperate zone.
We began the unit with Darwin's model of evolution:
Sources of variation Agents of change
unknown
Natural selection.
As a consequence of our modern understanding of heredity and genetics, we have
learned quite a bit about variation AND evolution. Our model, a this point in
the class is: Sources of variation Agents of Change
MUTATION:
-New Genes:
Natural Selection
point mutation
Mutation (polyploidy can make new species)
exon shuffling
RECOMBINATION:
- New Genes:
crossing over
-New Genotypes:
-crossing over
- independent assortment
What to know:
- Know the four levels at which a mutatin can occur, know how these occur, and know an example for each.
- Know why mutations are usually bad ('deleterious').
- How are new genes produced? List two ways (in eukaryotes).
Study Questions:
- Why are substitution mutations likely to have a less pronounced effect tha addition or deletion mutations?
- Outline the causes of genetic variation.
- How can polyploidy create a new species?
- Why are tetraploids more likely to establish a productive population than triploids?