Cell Biology
I. Overview
II. Membranes: How
Matter Get in and Out of Cells
III. Cellular Respiration
IV. Photosynthesis
V.
Protein Synthesis
VI.
Cellular Reproduction
Cell Biology
redux - why is this cell unit important, again?
This cell biology unit is getting
pretty long! We have discussed lots of very complicated processes at a molecular
and cellular level. It would be very easy to "lose sight of the forest
for the trees" - to become lost in the details of these processes and to
lose sight of their relevance to the evolutionary theme of the course. So, it
is a good time to put some things in our evolutionary context. The goal of this unit, in terms of the course's goal of understanding evolution, is to place DNA--the heritable, evolvable material--in it's functional context. What does it do? Well, it codes for proteins. Why is that important? Because proteins influence what gets into a cell, how that material is broken down for energy harvest, and how other proteins (and other molecules) are made. Proteins are the STUFF of life - they give 'shape' to life. Dna determines protein structure and function, so changes in DNA result in changes to the functions and 'shapes' of living things.... ie., changes to DNA are the cause of evolution.
We are using this cell unit as a
bridge to the topic of heredity.
Now we all know that the hereditary information is DNA, and that organisms like
bacteria, oak trees and house cats differ in their physiology, morphology, and
behavior because of differences in their DNA. In the heredity and population
genetics units, we will see how these differences are perpetuated through time,
and how changes in the genetic structure of populations takes place (evolution).
This cell unit is supposed to provide
you with a firm foundation regarding what DNA IS and what it DOES.
Only then can you truly appreciate why differences in DNA cause some
of the variation we see between cells, tissues, and multicellular organisms.
We have seen that proteins are instrumental in nearly everything that cells
- and therefore tissues, organs, and living organisms - do. Specific proteins
regulate the flow of material into and out of the cell (protein channels). Specific
protein catalysts (enzymes) co-ordinate the chemical reactions in the cell;
reactions that break down food (respiration) and harvest light energy (photosynthesis),
and reactions that build all the organic molecules needed in a particular cell
(like proteins in protein synthesis). Proteins are important structural components
of cells and organisms, too; like the actin in the cytoskeleton, the actin and
myosin in contractile muscle cells, and the collagen in skin and bone tissue.
And of course, proteins (histones and transcription factors) regulate DNA activity
- influencing which genes are transcribed in a given cell and when, and thus
determining which proteins are produced in a given cell and when. The functions
these proteins play is determined, in part, by their primary structure - their
sequence of amino acids. Because the sequence of nitrogenous bases in DNA is
the primary determinant of the sequence of amino acids in a protein (as we saw
in protein synthesis), it is the DNA sequence that is ultimately responsible
for what proteins do and what cells do. So, this cell unit was supposed to provide
you with an appreciation for how a cell functions and how the genetic system
controls this activity through the production of proteins.
So, THIS IS WHY our understanding
of DNA structure, function, and heredity are so important to evolutionary theory.
Changes in this heritable information cause changes in the physiological, morphological,
and behavioral characteristics of organisms. HOW do changes in the DNA cause
these changes in the physiology, morphology, and behavior of organisms? Hopefully,
the previous paragraph answered that question for you - by changing the proteins
that are responsible for cell, organ, tissue, and organism structure, function,
and development. You can only really understand this if you understand what
proteins do in a cell. And that is why cell biology is important to our understanding
of evolution; it is where DNA does its work, and it is where evolutionary, genetic
change has its most immediate biological effect.
Overview:
1. Why reproduce?
 Living
systems reproduce. In many ways, reproduction seems like the most purposeful
thing that living systems do. Indeed, most nature shows describe this attribute
as a "desire", "goal" or "urge", often described
in these same shows as a process performed "in order to perpetuate the
species". Well, it is currently impossible for us to ascertain the "desires",
"goals" or "urges" of an ameoba or an oak tree; or whether
the amoeba or oak tree is 'thinking' about the survival of its species as it
reproduces. Thankfully, Darwin's theory of natural selection absolves us from
having to understand "desires" - it explains the existence of complex
physiology, morphology, and behavior as a function of the relative benefit of
that trait to relative reproductive success.
Living
systems reproduce. In many ways, reproduction seems like the most purposeful
thing that living systems do. Indeed, most nature shows describe this attribute
as a "desire", "goal" or "urge", often described
in these same shows as a process performed "in order to perpetuate the
species". Well, it is currently impossible for us to ascertain the "desires",
"goals" or "urges" of an ameoba or an oak tree; or whether
the amoeba or oak tree is 'thinking' about the survival of its species as it
reproduces. Thankfully, Darwin's theory of natural selection absolves us from
having to understand "desires" - it explains the existence of complex
physiology, morphology, and behavior as a function of the relative benefit of
that trait to relative reproductive success.
In this context, the adaptive value
of reproduction is as obvious as the the difference between "1" and
"0". Think about it this way: the natural world is a dangerous place.
It is exciting and fun for a while, but all living things will eventually die
as a consequence of encountering an environment in which they cannot survive
(flood, fire, heat, or cold), or being eaten by a predator, or infected by a
pathogen, or simply by accident. So, the only life forms that will persist through
time are those that copy themselves at a faster rate than they are dieing. This
works from the cell level through the populational level, and even at the phylogenetic
level with respect to the persistence of particular lineages through geologic
time. So, for any population, if the birth rate remains lower than the death
rate then population will eventually go extinct. In a multicellular organism,
if the rate of cell production is lower that the rate of cell death, the organism
will waste away, losing tissue mass. At a geologic scale, lineages that produce
species faster than the extinction rate will persist longer through time that
lineages where the rate of speciation is lower than the rate of extinction.
So today, when we look at the entire diversity of the living world, we only
see descendants of those life forms that reproduced. And these living life forms
have inherited this capacity to reproduce, as well.
In terms of natural selection, members
of a population that do not reproduce at all have a differential reproductive
success of "0". Selection will favor organisms that evolve the capacity
to reproduce. For prokaryotes, cell reproduction occurs by binary fission. For
eukaryotic cells, cell reproduction occurs by mitosis. In single-celled protists,
mitosis produces two new organisms. In multicellular organisms, mitosis produces
new cells that can replace dead cells or increase the number of cells in the
organism. If the net number of cells increases, the multicellular organism grows.
As we have mentioned before, growth is usually a good thing. First, the bigger
you are, the fewer things can eat you. Second, becoming larger through multicellularity
allows for the increased efficiency and functional diversity of cell specialization.
2. An overview
of cell division:
Cell division is the process of producing
two functional 'daughter' cells from one ancestral 'parental' cell. In order
for both of the daughter cells to have the full functional repertoire of the
original parental cell, they must be able to make the full complement of proteins
that the parent cell makes. In order for this to happen, they must both receive
the full complement of genetic information (DNA) in the parental cell. Hmmm....
how can they BOTH get the FULL COMPLEMENT of genetic information in the parental
cell? Well, in order for this to happen, the parental cell must duplicate its
DNA prior to cell division. This process of DNA replication produces two full
complements of genetic information. Then, this genetic information must be divided
evenly, in an organized manner, to insure that both daughter cells get the complete
complement of information (and not a duplication of some information or an omission
of other information). Cells that receive an incomplete complement of genetic
information will not be able to make all the proteins the parental cell made,
and may not be able to survive. So, again, DNA replication and the process of
mitosis are of great selective, adaptive value. Only cells that replicate and
divide their genetic information evenly, with only minor errors or inconsistencies,
will be likely to survive. These survivors will pass on the tendancy to replicate
and divide their genetic information evenly, as well. So, there is very strong
selection ( a very large selective advantage) for correct DNA replication and
equal chromosomal allocation during mitosis.
These processes of DNA replication
and mitosis are only two stages in the life of a cell. To place them in context,
it's useful to consider the full life of a cell, from it's production by the
division of its parental cell through to its own division.
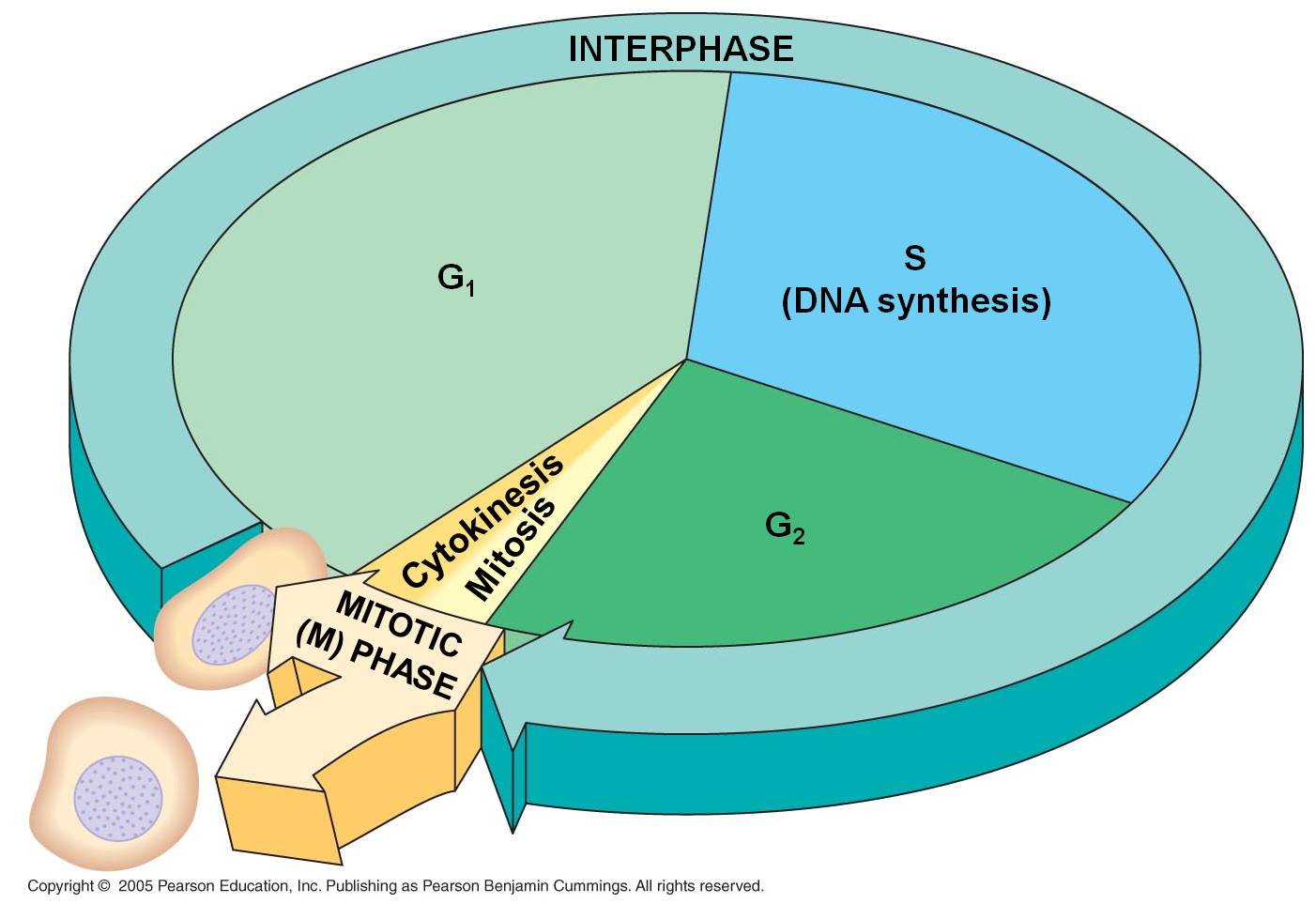 A.
The Cell Cycle
A.
The Cell Cycle
1. Interphase
- the 'interval' between divisions
a. G1
Our cell's life begins. That's sort
of a funny way to put it, because it seems to suggest that it is something new;
yet all of its constituents were part of the original parental cell. It is more
truly "1/2 an old cell with a full complement of DNA". Nevertheless,
it is an independent entity. In most protists, binary fission of the mitochondria
and chloroplasts occurs concurrently with the division of the nucleus during
mitosis, so the daughter cells have 'new' organelles, too. But in most multicellular
organisms, the allocation of organelles is largely a random process based on
how they are distributed in the cytoplasm during division. Then, the organelles
divide and 'repopulate' each daughter cell in G1.
The cell is roughly 1/2 the size
of the original parental cell. To grow to its appropriate size, it must synthesize
new biological molecules - and that means making the enzymes that will catalyze
those reactions. So, the DNA unwinds to the 'beads on a string' level, and the
genes between histones are available for transcription. When the DNA is unwound
('diffuse'), separate chromosomes cannot be seen with a light microscope. Rather,
the nucleus stains a uniform color except for one or several dark regions called
'nucleoli' (singular = nucleolus). These are areas were large amounts of r-RNA
are being synthesized and complexed with ribosomal proteins into functional
ribosomes. The ribosomes are exported from the nucleus to the cytoplasm, where
they will anchor to endoplasmic reticulum or the cytoskeleton.
Indeed, the G1 phase of a cell's
life is the most metabolically active period of it's life. It is growing in
size, and producing the proteins appropriate for its tissue type. Most cells
in multicellular organisms specialize during this period. Cells with very specific
structural adaptations to their specialized tissue type - like neurons with
long axons and muscle cells crammed with linear microfilaments - often remain
stalled in this stage after they become specialized; they do not divide again.
In this case, this stalled 'permanent' G1 phase is referred to a G0 ("G-nought').
 b.
S
b.
S
The S phase of the cell cycle is
when DNA replication occurs. The chromosomes are diffuse during this stage,
as well, so the enzymes (DNA polymerases) that replicate the DNA can access
the helices. Each double helix is separated, and the single strands are used
as templates for the formation of new helices on each template - changing one
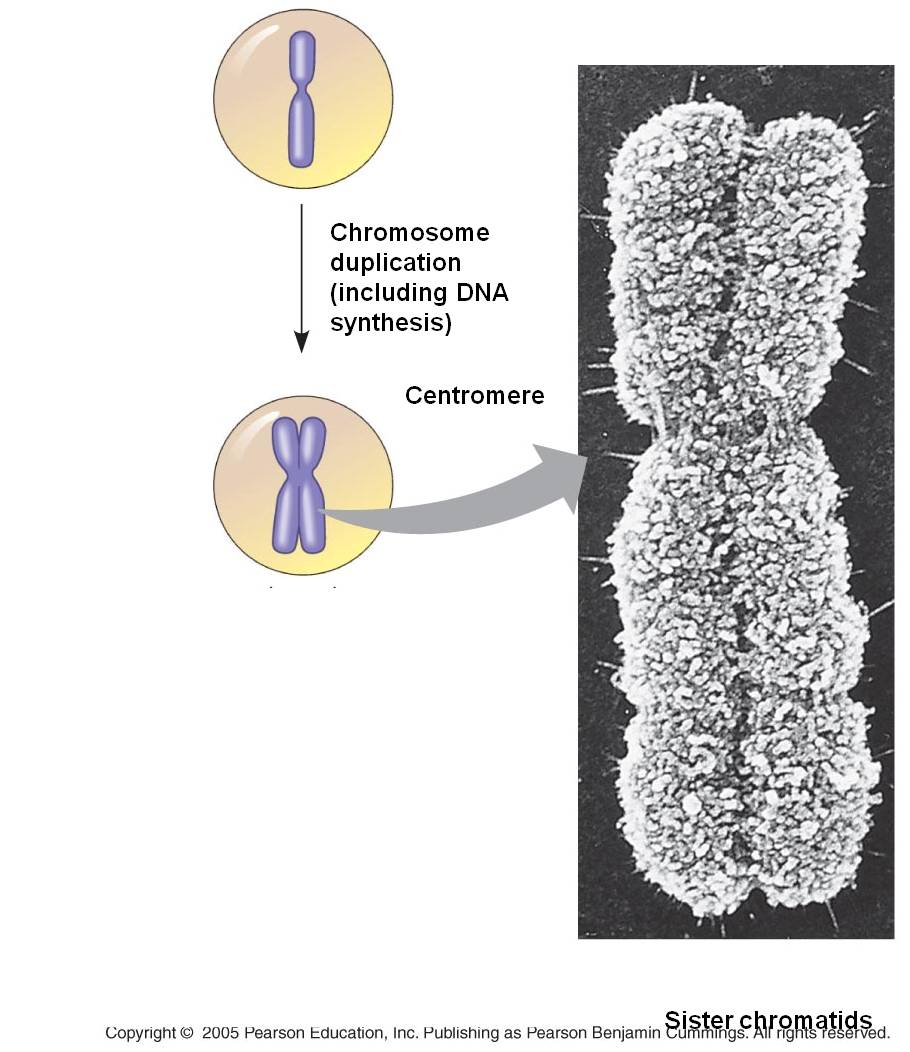
double helix into two. Terminology becomes a bit ambiguous here. A DNA double
helix is equivalent to a "chromatid". A chromosome may have one chromatid
(in its unreplicated form) or two chromatids (in its replicated form). DNA replication
is a rather complicated process described in more detail below. The transition
from the G1 to the S phase is a very critical stage in a cell's life cycle,
signalling the cell's progression towards division. In eukaryotes it is called
a 'restriction point'. Once the S phase begins, the cell will proceed through
to mitosis. This transition is orchestrated by a complex interplay of transcription
factors that regulate the activity of "cell division cycle genes".
These genes produce cyclin proteins that vary in concentration through the cell
cycle. They bind with 'cyclin-dependent kinases' and these cdk-cyclin complexes
activate transcription factors that initiate the next phase of the cell cycle.
The timing of the G1/S transition
is very important. During the G1 phase, the DNA is 'checked' by repair enzymes...
mismatched bases and other mutations are corrected. It is important that the
G1 lasts long enough for DNA repair to take place; otherwise any errors will
be copied during DNA replication and mutations will be passed to the next generation
of cells. There are several proteins that inhibit the progression of the cell
cycle - the most notable is called p53. This protein is a cell cycle inhibitor,
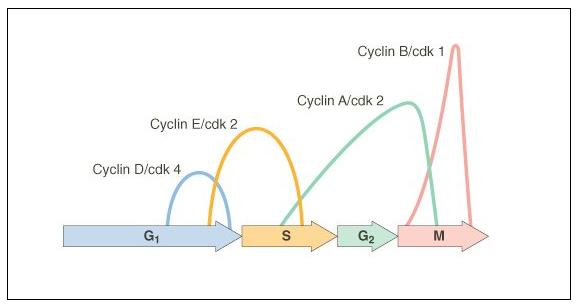 indirectly causing
the inactivation of cdk-cyclin complexes that would stimulate the onset of the
S phase. Mutations in this gene can make the protein non-functional; so cdk-cyclins
are not inhibited, and the onset of S happens quickly and prematurely - before
DNA repair is completed. This mutation is passed to the daughter cells, too,
along with all the other uncorrected mutations. These mutations accumulate with
each generation of cell division, affecting other genes that influence cell
function and specialization. This unregulated division of undifferentiated cells
creates a cancerous tumour. There are several other 'tumor suppressor' genes,
but mutations in p53 occur in 70% of small cell lung cancers, 80% of non-melanoma
skin cancers, and 60% of colon cancers. Obviously, correct regulation of the
cell cycle is critical to correct cell function and maintaining the integrity
of DNA.
indirectly causing
the inactivation of cdk-cyclin complexes that would stimulate the onset of the
S phase. Mutations in this gene can make the protein non-functional; so cdk-cyclins
are not inhibited, and the onset of S happens quickly and prematurely - before
DNA repair is completed. This mutation is passed to the daughter cells, too,
along with all the other uncorrected mutations. These mutations accumulate with
each generation of cell division, affecting other genes that influence cell
function and specialization. This unregulated division of undifferentiated cells
creates a cancerous tumour. There are several other 'tumor suppressor' genes,
but mutations in p53 occur in 70% of small cell lung cancers, 80% of non-melanoma
skin cancers, and 60% of colon cancers. Obviously, correct regulation of the
cell cycle is critical to correct cell function and maintaining the integrity
of DNA.
c. G2
After DNA replication is complete
the cell goes through another rapid period of growth in preparation for mitosis.
The DNA is checked again for damage caused and errors made during DNA replication.
Once again, p53 inhibits the transition to the mitotic phase, providing time
for this repair to take place. In cancer cells with mutations in p53, the G2
phase may be nearly eliminated, with the cell proceeding directly from DNA replication
to mitosis. CDK's bind to new cyclins, and these complexes active a different
set of proteins that initiate mitosis.
2. Mitosis
The process of mitosis can be summarized
as follows: the chromosomes condense, making it easier to divy them up evenly.
The replicated chromsomes are aligned in the middle of the cell by cytoskeletal
fibers. Each chromosome consistes of two identical double helices, called chromatids.
During the process of mitosis, these chromatids separate from each other, and
one double-helix from each chromosome is pulled to each end of the cell. The
membrane and cytoplasm are divided and the nuclear membrane reforms around the
chromosomes in each daughter cell. We will look at this process in more detail,
below.
 B.
Mitosis
B.
Mitosis
Mitosis is a continuous process of
chromosome condensation, chromatid separation, and cytoplasmic division. This
process is punctuated by particular events that are used to demarcate specific
stages. This process was first described by Walther Flemming in 1878, we he
developed new dyes and saw 'colored bodies' (chromo-somes) condensing and changing
position in dividing cells. He also coined the term 'mitosis' - the greek word
for thread - in honor of these thread-like structures.
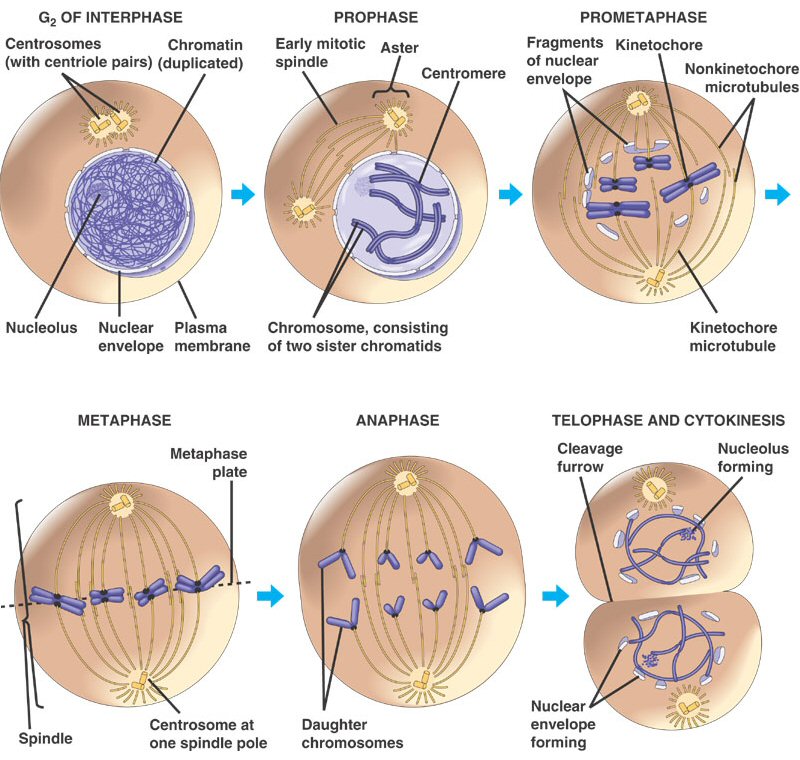
1. Prophase:
The transition from G2 to Prophase of Mitosis is marked by the condensation
of chromosomes.
2. Prometaphase:
The chromosomes continue to condense, and the nuclear membrane disassembles.
The microfibers of the spindle apparatus attach to the kinetochores on the replicated
chromsomes.
3. Metaphase:
The spindle aranges the chromosomes in the middle of the cell.
4. Anaphase:
The proteins gluing sister chromatids together are metabolized, and the sister
chromatids are pulled by their spindle fibers to opposite poles of the cell.
It is important to appreciate that these separated chromatids (now individual,
unreplicated chromosomes) are idnetical to one another and identical to the
orignial parental chromosome (aside from unrepaired mutations).
5. Telophase:
The cell continues to elongate, with a concentrated set of chromosomes at each
end. Nuclear membranes reform around each set of chromosomes, and the chromosomes
begin to decondense.
6. Cytokinesis:
Cytokinesis is sometimes
considered a part of telophase. In this stage, the cytoplasm divides. In animal
cells, the membrane constricts along the cell's equator, causing a depression
or cleavage around the mid-line of the cell. This cleavage deepens until the
cells are pinched apart. In plants, vesicles from the golgi coalesce in the
middle of the cell, expanding to form a partition that divides the cell and
acts as a template for the deposition of lignin and cellulose that will form
the new cell wall between the cells.
As a consequence of this process,
two cells are produced from one parental cell, each having a complete complement
of genetic information - a copy of each original chromosome that was present
in the parental cell. Each of these cells now begins the G1 phase of interphase.
Things to KNOW:
1. Draw
the cell cycle, labeling each stage and highlighting the main event in each
stage.
2. Draw a
chromsome before and after replication; both at the molecule level (showing sequences of A, T, G, C...) and the chromosomal level (use the terms chromosome and chromatid).
Study Question:
3. Draw a
cell, 2n = 6, and show each of the stages of mitosis. Write a brief description
of the events of each stage.