 A.
Pre-Mendelian Ideas about Heredity:
A.
Pre-Mendelian Ideas about Heredity:
In this unit, we will examine how DNA is transferred through time, we will describe the process of sexual reproduction, and we will solve Darwin's Dilemma regarding the source of heritable variation.
A.
Pre-Mendelian Ideas about Heredity:It was common knowledge that offspring looked alot like their parents. This observation was especially obvious and important in the agrarian societies of the pre-industrial age, where people saw that certain traits in domesticated plants, domesticated animals, and humans 'ran in families'. In fact, this was not just an idle issue. Understanding how heredity worked was an important economic question to these farmers, who were trying to breed more valuable and productive crops and livestock.
1. Preformationist Ideas
 One
school of thought - the "preformationist" school, thought that a miniature
human lived within either the egg (the 'ovist' school), or the sperm ('the homunculan'
school). This offspring laid in a state of suspended animation until the sperm
stimulated the egg to grow (for the ovist school), or the sperm was placed in
the fertile womb to grow ('homunculan' school). Of course, both of these ideas
had serious flaws. If the offspring was really just a product of one sex, then
why did the offspring have characteristics of both parents? Some theologians
saw interesting implications, too. If all generations were preformed, then each
generation must be nested within the egg or sperm of the preceeding generation
- also already preformed. The generations of humans would be like a set of russian
dolls, one nested with the other. At some point, it would seem that nature would
reach the limit of how small a preformed person could be - and that would be
the last generation (which would be followed by the apocalypse).
One
school of thought - the "preformationist" school, thought that a miniature
human lived within either the egg (the 'ovist' school), or the sperm ('the homunculan'
school). This offspring laid in a state of suspended animation until the sperm
stimulated the egg to grow (for the ovist school), or the sperm was placed in
the fertile womb to grow ('homunculan' school). Of course, both of these ideas
had serious flaws. If the offspring was really just a product of one sex, then
why did the offspring have characteristics of both parents? Some theologians
saw interesting implications, too. If all generations were preformed, then each
generation must be nested within the egg or sperm of the preceeding generation
- also already preformed. The generations of humans would be like a set of russian
dolls, one nested with the other. At some point, it would seem that nature would
reach the limit of how small a preformed person could be - and that would be
the last generation (which would be followed by the apocalypse).
2. Epigenetic Ideas
The other major school of thought promoted the "epigenetic" idea. Here, the egg does not actually contain a small individual, but rather only the potential to grow and develop into an individual. This idea had much less support, because there were no ideas about how this could happen; the growth of a small human was alot easier to understand than the development of organs and organ systems from 'nothing' - or at least, from things that weren't organs or organ systems! However, the epigenetic idea had one major benefit - it could explain why offspring often expressed characteristics of both parents. If both parents contributed 'stuff' to this epigenetic mass of potential, then the developing offspring could inherit traits from both parents.
There was an additional observation that was very problematic with this blending idea, however. Most people were well aware that some traits 'skipped a generation', or even many generations. The reappearance of an ancestral trait (" you have your great-grandmother's nose") was know as a 'sport'. If the hereditary information from two parents were fluids that mixed, like black and white paint, it seemed difficult to explain how, after mixing black and white into grey, you could get some pure white paint back out of the mixture in a later generation.
3. Blending
Because semen is a fluid, it seemed likely that the genetic information was a fluid. The 'mixing' of the heredity fluids from two parents might explain, sort of, why the offspring expressed a mixture of traits from both parents. . Mendel
set out to test these alternate hypotheses by breeding organisms from different
'pure-breeding' stocks together. In other words, Mendel conducted hybridization
experiments, forming hybrids by crossing strains that bred true for particular
characteristics. He selected the common garden pea, Pisum sativum,
as his study organism. He chose this plant because there were alot of pure-breeding
varieties already available; farmers had selected for different characteristics,
creating strains that produced round peas, wrinkled peas, yellow peas, green
peas, tall plants, short plants, etc. Indeed, Mendel reports that he received
34 different strains from farmers. In addition, because peas were fast growing
and fecund (produced lots of offspring - each pea is a separate offspring),
he could generate a lot of data in a matter of months. (Some characters, however,
like flower color or plant height, could only be determined in the following
year, when the offspring grew up and exhibited the trait.) Being trained in
mathematics and probability, he knew that a large sample would provide a 'truer'
indication of a pattern than a small sample.
Modern geneticists have chosen other study organisms - like fruit flies, mice,
and bacteria - for the same reason.
Mendel
set out to test these alternate hypotheses by breeding organisms from different
'pure-breeding' stocks together. In other words, Mendel conducted hybridization
experiments, forming hybrids by crossing strains that bred true for particular
characteristics. He selected the common garden pea, Pisum sativum,
as his study organism. He chose this plant because there were alot of pure-breeding
varieties already available; farmers had selected for different characteristics,
creating strains that produced round peas, wrinkled peas, yellow peas, green
peas, tall plants, short plants, etc. Indeed, Mendel reports that he received
34 different strains from farmers. In addition, because peas were fast growing
and fecund (produced lots of offspring - each pea is a separate offspring),
he could generate a lot of data in a matter of months. (Some characters, however,
like flower color or plant height, could only be determined in the following
year, when the offspring grew up and exhibited the trait.) Being trained in
mathematics and probability, he knew that a large sample would provide a 'truer'
indication of a pattern than a small sample.
Modern geneticists have chosen other study organisms - like fruit flies, mice,
and bacteria - for the same reason.
1. Monohybrid Experiments (single traits)
Mendel reported the results of seven parallel experiments. In each experiment, he cross plants that had different expressions of a particular trait. So, in the third set of trials, he bred plants that 'bred true' for white flowers (when bred with themselves) with plants that produced purple flowers. For this experiment, he did 35 cross-fertilizations with ten plants.
a. Reciprocal parental crosses tested the ovist and homunculan schools:
A very important methodological aspect of the experiment was that he conducted reciprocal crosses between stocks. So, he placed pollen pure-breeding white flowers on the stigma of purple-flowers, and pollen from purple flowers on the stigmas of white flowers. So, by comparing the results of these reciprocal crosses, he could see if the inheritance of the trait (flower color) was associated with the sex of the parent. If the homunculan school was correct, then the offspring should bear the trait expressed in the stock that donated the pollen. If the ovist school was correct, then the offspring should bear flowers the color of the strain that received the pollen and donated the egg (in the ovule).
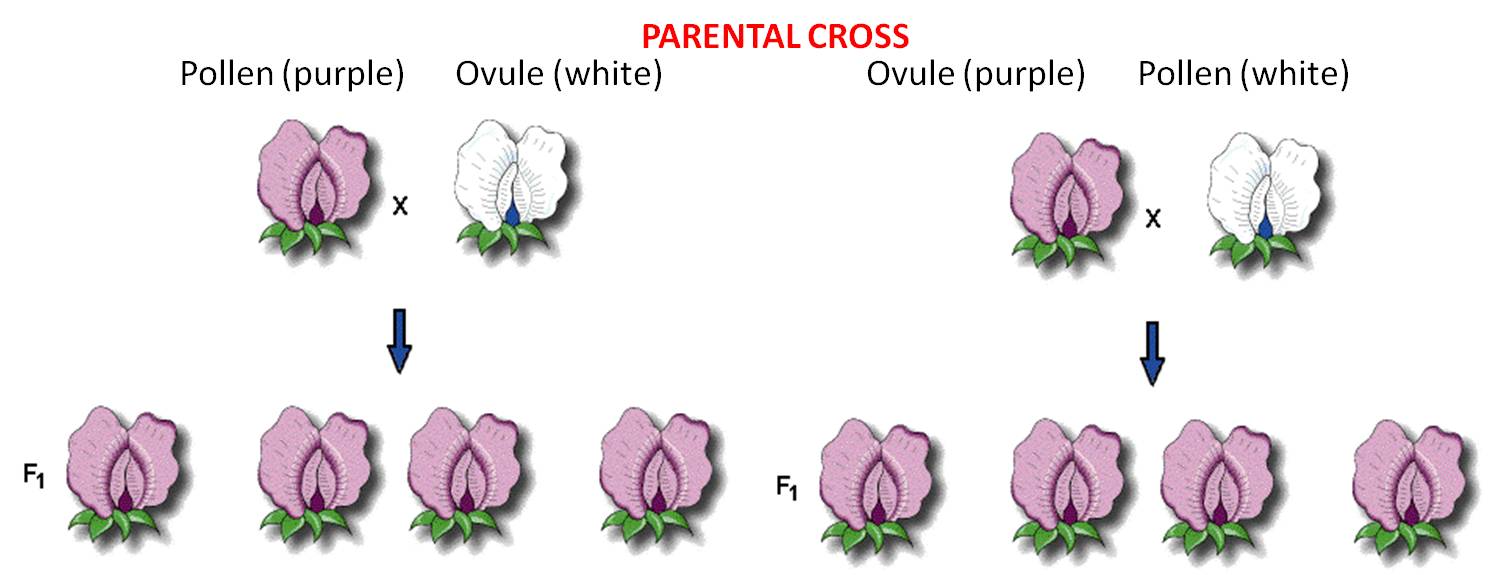 These 'pure-breeding'
stocks were called the 'parental generation'.
These 'pure-breeding'
stocks were called the 'parental generation'.
pollen (male) from Purple flowers X ovule (female) from White flowers
AND
pollen (male) from White flowers X ovule (female) of Purple flowers
These crosses produced offspring (peas). The first generation of 'hybrids' produced from pure-breeding parental strains is called the F1 (for 'first 'filial') generation. He planted and grew the peas produced from both crosses and observed their flower color. In both reciprocal crosses, all the plants produced purple flowers. These results falsified both the homunculan and ovist schools; both hypotheses predicted that plants from one or the other cross would yield white flowers. Indeed, Mendel realized that these results suggested that both parents had to contribute hereditary factors for this trait; the purple offspring from the first reciprocal demonstrated that the male contributes, and the purple offspring in the other reciprocal demonstrated that the female contributes, too. But if both parents contributed hereditary information, what happened to the white information contributed by the other parent in each cross? Mendel hypothesized that the contribution was present in the offspring, but not expressed. He coined the term 'dominant' for the trait expressed in the F1, and 'recessive' for the trait he thought was there, but hidden. He became more confident in these hypotheses when all seven traits that he studied showed the same pattern.
 b.
Crossing the F1 hybrids tested the blending hypothesis:
b.
Crossing the F1 hybrids tested the blending hypothesis:
He allowed the F1 plants to fertilize themselves, essentially performing an F1 x F1 cross. He makes reference to the fact that, in many species and for many traits, the hybrids (F1's) exhibit a triat intermediate to the two parental types. This intermediate result might be expected of a blending pattern (white paint and black paint making grey), but one trait dominating the other could also be produced from a blending mechanism, too. One thing you would not expect from a blending of fluids, however, would be the re-expression of the recessive 'white' trait. Grey paint mixed with grey paint should not produce white paint.
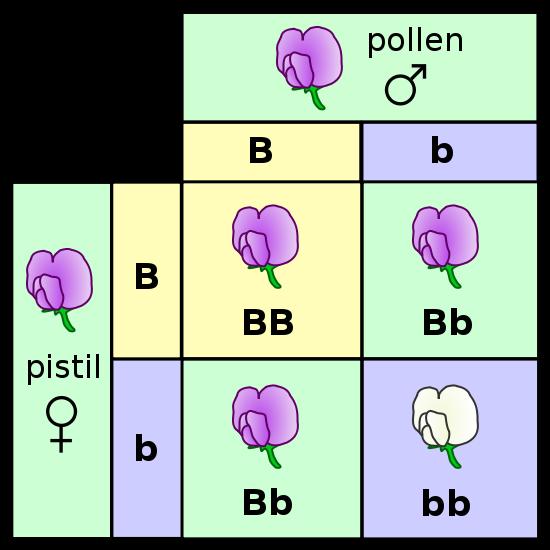
The peas from these crosses were grown and the flowers were observed. Of the 929 plants that he grew from these seeds, 705 bore violet flowers and 224 had white flowers; a ratio of 3.15:1. In fact, all seven traits produced ratios near a 3:1 ratio in the F2 generation (offspring of F1 x F1 crosses). He recognized the importance of the 3:1 ratio when combining two things. It reminded him of a simple binomial expansion (a + b)2 = 1aa + 2ab + 1bb. If an organism has 2 different types of hereditary units (a = purple and b = white) for a given trait and it mates with itself, then there are three combinations that are possible in the offspring (aa, ab, bb). And, they should occur in a 1/4:2/4:1/4 ratio. If, as a consequence of dominance, the progeny with different particles (ab) only express one - the 'a' (purple) - then they will look like the aa's and 'a' (purple) offspring will occur 3/4 of the time and 'b' (white) offspring will occur only 1/4 of the time.
c. Mendel proposed four postulates (hypotheses) to explain these data:
- genetic info is unitary or 'particulate' - it is not a fluid that blends
- each oganism has two unit factors (we call these 'genes' now) for each trait
- if an organism has different factors for a given trait, one is expressed (dominant) over the other (recessive)
- the two genes for a given trait separate and go into separate gametes during gamete formation. Subsequent fertilization (fusion of gametes) is random. This is called Mendels First Principle: Principle of Segregation
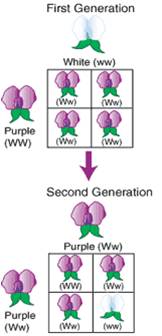 d.
How his hypothesis explains his observations:
d.
How his hypothesis explains his observations:
Parentals: WW (Purple) x ww (white) .... (and the reciprocal)
Gametes: W w
F1 offspring:
Genotypic Ratio: 100% Ww (heterozygous)
Phenotypic Ratio: 100% Purple
F1 x F1 Cross: Ww x Ww
Gametes:
W and w W
and w
F2 offspring:
Genotypic Ratio: 1/4 WW, 2/4 Ww, 1/4 ww Some Terms:
Phenotypic Ratio: 3/4 Purple, 1/4 White
Genotype = the type of genes an organism has, like Ww.
Genotypic Ratio = is the fractional representation of different
genotypes in a group, such as 1/4WW : 1/2Ww : 1/4 ww
Phenotype = the characteristic or trait that is expressed,
such as 'Purple'. Usually, this is represented by the gene that causes the trait,
such as 'W' in the example, above.
Phenotypic Ratio = the fractional representation of different
phentypes in a group, such as 3/4W : 1/4 w.
2. Monohybrid Test Crosses
If the hypotheses are correct, Mendel reasoned, then the F1 has a gene for the recessive 'white' trait that is not expressed. How could he determine whether it was there or not? He realized that he could answer this question if he crossed it to the recessive white parent: (ww). The logic is this: the homozygous recessive parent can only pass recessive alleles to the offspring. Whatever the offspring receives from the OTHER parent will determine the expression of the trait. It the offspring receives a dominant allele for purple from the other parent, then the offspring (Ww) will express purple flowers. But, if the other parent is hiding a recessive allele and passes that to the offspring, then the offspring will have the homozygous recessive genotype (ww) and will produce white flowers.
Mendel predicted that, under this hypothesis, the phenotypic ratio in the offspring would be: 50% purple (W): 50% white (w). The results supported his hypothesis, and supported his postulates. Replication of this results across all seven traits gave strong confirmation that he had described a general pattern of heredity.
More
Terms:
Homozygous: A zygote is a fertilized egg that has received
genes from both parents. If the genes received from both parents for a given
trait are the same, then this zygote (and the organism that develops from it)
is homozygous for this trait. If the zygote has received dominant genes
for the trait from both parents, it is homozygous dominant (WW); if
it has received the recessive allele from both parents, it is homozygous
recessive (ww).
Heterozygous: If the zygote receives different alleles from
the parents for a given trait, then the zygote is heterozygous for
that trait (Ww).
Alleles are different 'forms' of a gene, that affect a given
trait - like flower color - in different ways ('W'and 'w' are alleles for flower
color).
3. Dihybrid Experiments
Most animal and plant breeders had noticed that some traits seem to be inherited together. Indeed, in his own experiments, Mendel saw that flower color was always correlated with the color of the seed coat, such that seeds with white seed coats grew into plants with white flowers, and seed with brown seed coats grew into plants with purple flowers. Having demonstrated that his seven traits all exhibited the same pattern of inheritance, he now set out to determine whether these traits were inherited in a dependent or independent manner. This is a very insightful question. For example, he had already demonstrated that yellow pea color was dominant to green, that round pea shape was dominant to wrinkled, and that F1xF1 crosses for each trait produced a 3:1 ratio in the offspring. Well, maybe these traits are inherited in the same way because they are governed by the SAME heritable unit factor (gene)... and that's why they produce the same pattern. If yellow and round were caused by the same gene, then of course they would be inherited in the same way. Or, maybe they are governed by different unit factors that have a pattern of heredity that is related to one another... maybe they are governed by different factors that "travel" from parent to offspring together - yellow always traveling with round, for instance. This had important practical applications, too. A sheep farmer would profit from sheep that produce both good wool and good milk (for feta or roquefort cheese) from his sheep herd. But, maybe the trait of quality wool is inherited with the trait of poor milk production. In that case, they might not want to strongly select for great wool quality, as milk production would suffer. It might be best to maintain a variable population that produces good wool and adequate milk. Every organism expresses a combination of traits, so addressing how combinations of genes are inherited is a very important question.
a. Parental crosses of stocks breeding true for two traits
First, Mendel bred plants together and created stocks that bred true for two traits: seed color and seed shape. He created a stock of plants that produced yellow, round peas and a stock that produced green, wrinkled peas. He already knew the dominance patterns, and had already concluded that parents bore two 'unit factors' (genes) for each trait. So, he hypothesized that the genotypes of the plants in each pure-breeding stock would be RRYY (round, yellow) and rryy (wrinkled green). He placed pollen from the wrinkled, green stock on flowers of the yellow, round stock and observed the progeny.
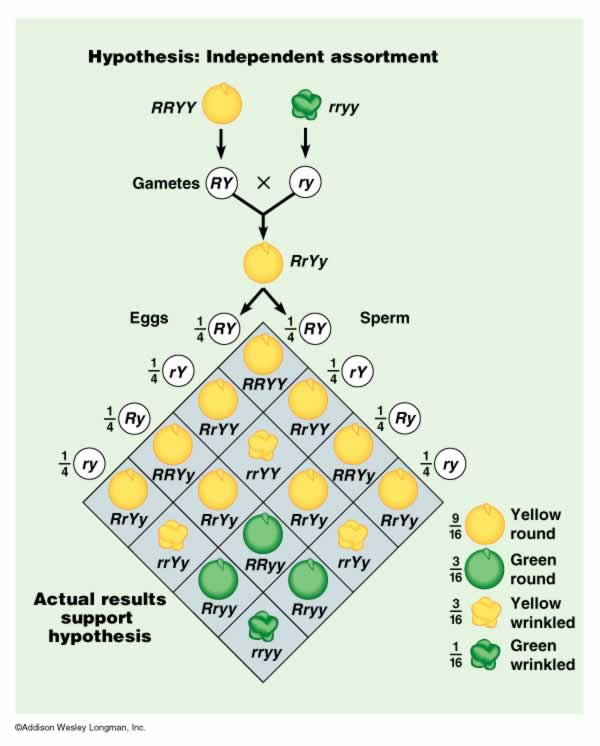 Round, Yellow (RRYY) x wrinkled, green (rryy)
Round, Yellow (RRYY) x wrinkled, green (rryy)
He hypothesized that the parental types would produce RY gametes and ry gametes, respectively. The combining of the genes in the offspring would make only RrYy genotypes, that should all express the phenotypic traits of yellow, round peas. This hypothesis was confirmed. However, this did not yet address the issue of independence; this pattern would also have occurred if color and shape were governed by one gene, or if the genes were inherited in a dependent manner. In short, the Y had to be inherited with an R, because that's all the RRYY parent had to give.
b. Crossing the F1's:
He crossed the F1 individuals, and found the following results:
- 315 round, yellow peas (~ 9/16)
- 101 wrinkled yellow peas (~3/16)
- 108 round green peas (~3/16)
- 32 wrinkled green peas (~1/16)
TOTAL - 556 peas
c. His interpretation and hypothesis:
First, it was obvious that the two traits were not caused by the same gene; all yellow peas were not ALSO round - he had produced some yellow wrinkled peas. So, different genes governed see color and seed shape, and so the genes for these traits could occur in differnt combinations.
Mendel realized that the 3:1 ratios were still preserved for each trait, when considered separately. For example, taken together, the ratio of yellow:green peas was 416:140 (2.97:1) and the ratio of round:wrinkled peas was 423:133 (3.18:1).
And also, Mendel realized that the
results of this experiment, and the results of subsequently self-crossing these
hybrids, were consistent with the probability of occurrence of independent events.
We know this as The Product Rule: "The probability that independent events
will occur together is the product of their independent frequencies."
These ratios in which the traits occurred in combination equaled the products of the independent frequencies:
3/4 W X 3/4 Y = 9/16 WY (round yellow)
1/4 w X 3/4 Y = 3/16 wY (wrinkled yellow)
3/4 W x 1/4 y = 3/16 Wy (round green)
1/4 w x 1/4 y = 1/16 wy (wrinkled green)
Mendel's hypothesis can be stated like this:
During gamete formation, the way one pair of genes (governing one trait) segregates is not affected by (is independent of) the pattern of segregation of other genes; subsequent fertilization is random. This is called Mendel's Second Principle: Principle of Independent Assortment.
He tested this hypothesis is subsequent experiments using three traits (seed color, seed shape, and seed coat color), and showed that the combination of traits expressed in the offspring were consistent with predicitons from their independent frequencies. For example, In a "trihybrid" F1 cross (a self-cross of an individual that is heterozygous for all three traits - YyRrCc), the fraction of peas that are yellow, round, and have a brown seed coat (caused by the dominant allele, "C") should be 3/4 x 3/4 x 3/4 = 27/64. His results confirmed his hypothesis.
4. Dihybrid test cross:
We can test for independence a bit more directly, through a dihybrid test cross. The principle of independent assortment is really predicting what the frequencies of different gamete types will be. So, consider an organism that is a double heterozygote (RrYy). The principle of independent assortment predicts that the segregation of the R genes (R and r) during gamete formation will be unrelated to (independent of) the pattern of segregation of the Y genes (Y and y) going into the same gametes.
With respect to the "R" gene, it has the Rr genotype and should produce R and r gametes at a ratio of 1/2 R to 1/2 r.
Likewise, with respect to the "Y" gene, the organism should produce Y and y gametes at a ratio of 1/2 Y to 1/2 y.
Now, every gamete must carry one (and only one) gene for each trait. So first of all, there can't be Rr or Yy gametes. And, under the hypothesis of independent assortment, the types and frequencies of the gene combinations that occur in the gametes can be predicted as the product of their independent frequencies. So, we would predict that this organism would produce 1/4 RY gametes (1/2 R x 1/2 Y), 1/4 Ry gametes (1/2 R x 1/2 y), 1/4 rY gametes (1/2 r x 1/2 Y), and 1/4 ry gametes (1/2 r x 1/2 y). These gametes are in the first column in the figure, below.
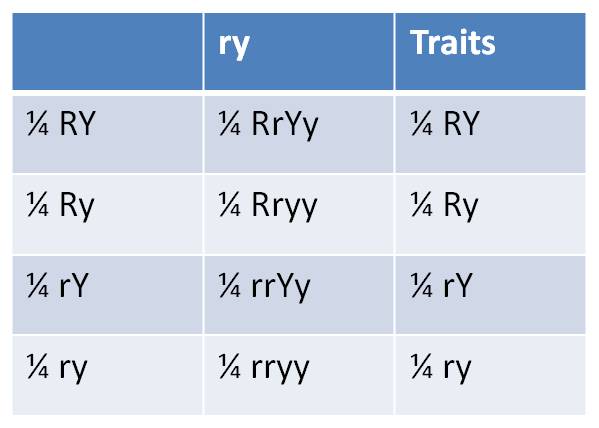 So,
to test independent assortment, we really want to know whether this organism
is producing gametes in these predicted frequencies. But we can't see the genes
in the gametes too easily; through heredity, all we can observe are the traits
in the phenotype of the offspring. And since these gametes carry recessive alleles,
it seems likely that they would be hidden from our observations.
So,
to test independent assortment, we really want to know whether this organism
is producing gametes in these predicted frequencies. But we can't see the genes
in the gametes too easily; through heredity, all we can observe are the traits
in the phenotype of the offspring. And since these gametes carry recessive alleles,
it seems likely that they would be hidden from our observations.
Unless we are clever.
If we mate this first organism to a second organism homozygous for the recessive traits (rryy), then the second parent will only donate recessive alleles (ry) to the offspring. (This gamete is in the header of the second column, at right). In this "test cross", the genes the offspring receives from the first parent will be expressed in the phenotype - we will be able to "see" the gene combinations, and their frequencies, produced in the gametes of the first individual. (The 'traits' expressed by the offspring - shown in the third column - are the same traits in the same frequencies as the gamete types produced by the first parent - shown in the first column.)
1. If the genes assort independently, then you can calculate ‘single gene’ outcomes and multiply results together…
For Example: AaBb x Aabb:
- what is the probability of an Aabb offspring?
- What is the probability of an offspring expressing Ab?
- How many genotypes are possible in the offspring?
- how many phenotypes are possible in the offspring?
Do the single-gene Punnett squares:
 and
and 
Answer each question for each gene, and then multiply the answers together:
- what is the probability of an Aabb
offspring? 1/2 (Aa) x 1/2 (bb) = 1/4
- What is the probability of an offspring expressing Ab? 3/4
(A) x 1/2 (b) = 3/8
- How many genotypes are possible in the offspring? 3
x 2 = 6
- how many phenotypes are possible in the offspring? 2
x 2 = 4
2. You can easily address more difficult multigene problems:
For Example: (female) AaBbCcdd x AABbccDD (male)
- how many types of gametes can each
parent produce?
- What is the probability of an offspring expressing ABCD?
- How many genotypes are possible in the offspring?
- how many phenotypes are possible in the offspring?
Answer the questions for each gene, then multiply the answers together:
- how many types of gametes can each parent produce?
| FEMALES - types of gametes at each gene (locus) | MALES - types of gametes at each gene (locus) |
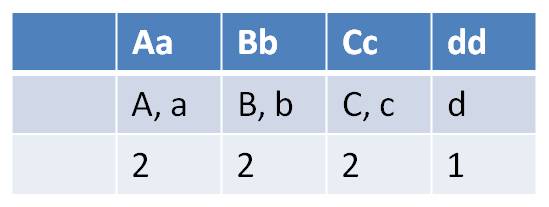 |
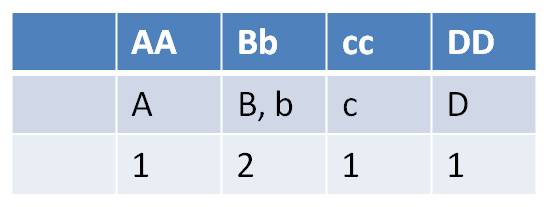 |
| Females can make 2 x 2 x 2 x 1 = 8 gamete types | Males can make 1 x 2 x 1 x 1 = 2 gametes types |
With respect to offspring, do the single-gene Punnett squares:
 |
 |
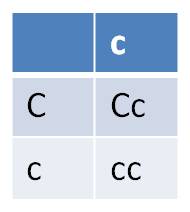 |
 |
- What is the probability of an offspring
expressing ABCD? = 1 (A) x 3/4 (B) x 1/2 (C) x
1 (D) = 3/8
- How many genotypes are possible in the offspring? =
2 x 3 x 2 x 1 = 12
- how many phenotypes are possible in the offspring? =
1 x 2 x 1 x 1 = 2
Note that for the C and D genes, we simplified the Punnett Squares. If a parent is homozygous for a gene, then it can only make one type of gamete with respect to that trait. So, to reduce redundancy, we just used one column for the 'cc' parent in the third Punnett Square, and only one row and one column for the parents in the last Punnett Square. The same could have been done for the AA parent in the first Punnett square - the information in the two columns is identical.
Meiosis produces specific types of reproductive cells. In animals, meiosis ONLY occurs in the gonads (ovaries/testes) and produces eggs or sperm. The production of egg and sperm occurs for only one purpose - sexual reproduction. Sperm and egg must fuse (fertilization) before the development of the offspring can begin. Spores do not fuse; a spore is a single reproductive cell that simply begins to divide and grow on its own to become an offspring. In this lecture, we will ask the question "why sex?" Of course, in an evolutionary context, this question means "is this complex trait adaptive, and if so, how?" After describing the adaptive significance of asexual and sexual reproduction, we will describe the process, its relevance to Mendel's principles of heredity, and we will solve Darwin's dilemma regarding the source of heritable variation.
1. How? - Reduction and Fertilization
There is a big benefit to combining genes from different organisms - new variations are produced that may be adaptive. Some organisms donate genes between cells, others trade nuclei. But for most sexually reproducing organisms, this mixing of genomes occurs through the production of specialized cells called gametes that fuse to form a new genotype and a new organism.
There is a problem to fusing normal body cells to produce this variation. By fusing body cells, the number of chromosomes and the amount of genetic information doubles each generation. As we'll see, many of the chemical reactions and patterns of genetic regulation depend on a constant balance of enzymes and substrates, and a constant balance of particular proteins. Doubling the genetic information can disturb these quantitative relationships. In addition, fusing body cells does not escape the constraints of Muller's ratchet - all genes are passed to offspring and the offspring now acquire all the bad genes from BOTH parents.
The solution that life evolved is a new process of cell division - meiosis. In this process that produces specialized reproductive cells, the genetic information is halved. This halving is also called reduction. If the cells produced by this process are gametes (remembering that plants and fungi use meiosis to produce spores), then the fertilization of sperm and egg reconstitutes the appropriate set of genetic information for that species.
Meiosis has two cycles of division, preceeded by an interphase containing a G1, S, and G2 phase. So, the cell entering meiosis has replicated chromosomes. The first cycle is called Meiosis I, or the reduction cycle. A diploid cell containing two sets of replicated chromosomes divides and produces cells that are haploid, containing one set of replicated chromosomes. The second cycle is Meiosis II, also called the division cycle. In this cycle, the haploid cells divide in a manner much like mitosis. In other words, the sister chromatids of each replicated chromosome are separated, but there is no change in ploidy. The haploid cell with replicated chromosomes divides into haploid cells with unreplicated chromosomes .
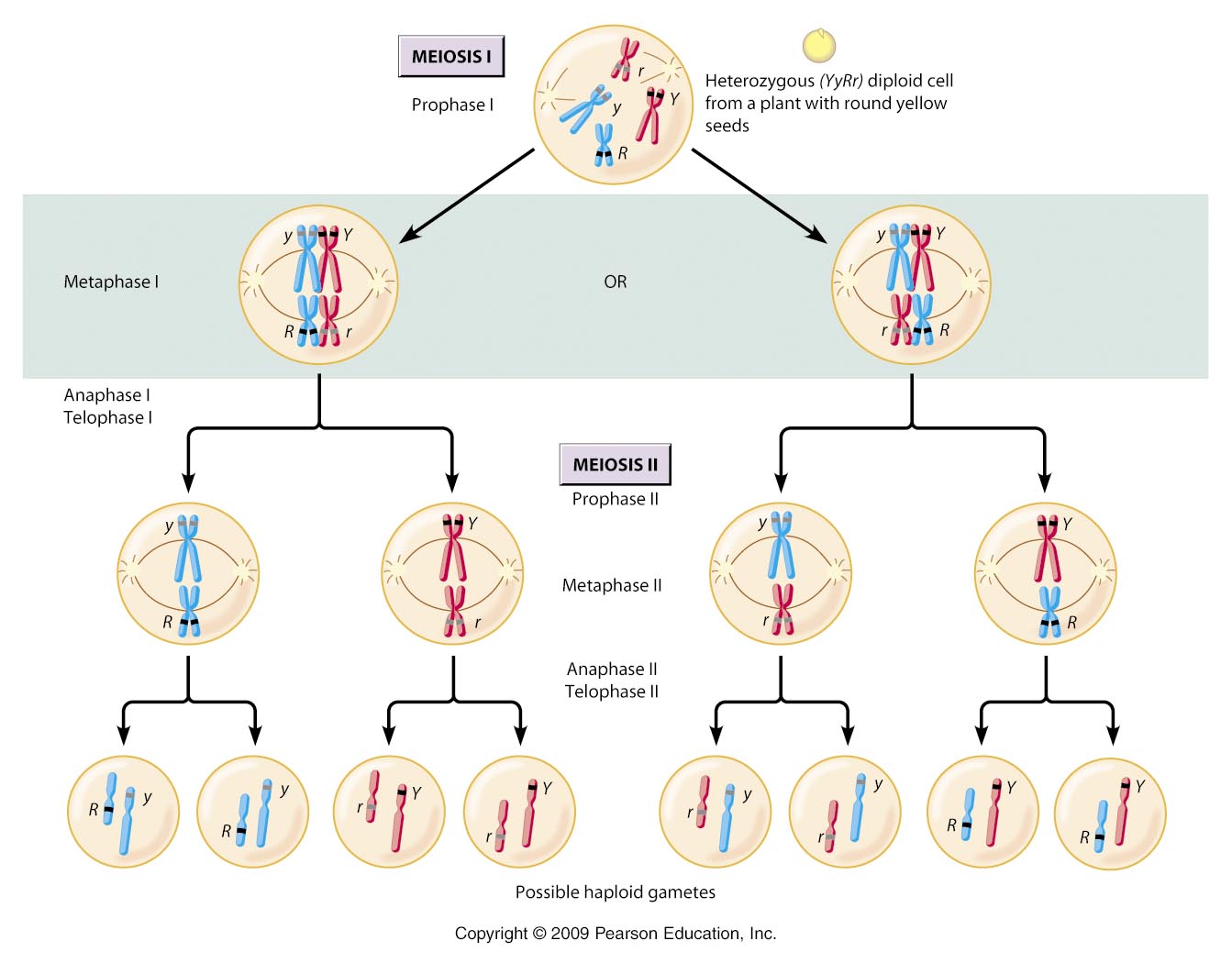 1.
Meiosis and Mendelian Heredity
1.
Meiosis and Mendelian Heredity
Meiosis was first described in 1876 by Oscar Hertwig; the movement of chromosomes in meiosis was first described by Eduard Van Beneden in 1888. Sutton and Boveri, working independently in 1902, saw pairs of chromosomes separating in Meiosis I. Being familiar with the recently rediscovered work of Gregor Mendel, they appreciated this correlation between the movement of chromosomes during gamete formation and Mendel's principle of segregation. If one homolog carried one allele for a trait while the other homolog carried the other allele, then the separation of homologs could explain the segregation of alleles during gamete formation. Likewise, if the way that one pair of homologs separated had no effect on the way that other pairs of homologs separated, then the movement of chromosomes could explain the principle of independent assortment, as well.
They hypothesized that these chromosomes, moving in a pattern consistent and correlated with Mendel's two principles, might contain the heredity determinants. This is a big deal - it is called the "Chromosomal theory of heredity". Why do we know that chromosomes carry the genetic info? Because in 1902, Sutton and Boveri observed the correlation between the movement of chromosomes and Mendel's laws of heredity, and they proposed the hypothesis that chromosomes might carry the heredity information and cause patterns of heredity. This hypothesis was confirmed by Thomas Hunt Morgan's group working with fruit flies in 1915. Experiments in 1944 determined that it was the nucleic acid in chromosomes, and not the proteins, that was the hereditary information.
2. Solving Darwin's Dilemma:
Darwin observed that every population expressed considerable variation in every generation. He was confused by this, because if selection had been operating for millennia, why hadn't selection honed a species to the point where only the best adapted phenotype persisted and reproduced? In addition, Darwin was a supporter of the idea of blending heredity. This would further reduce variation each generation (white and black to grey...). However, because the hereditary material is PARTICULATE and not a blending fluid, sexual reproduction produces unique combinations of particles in EVERY generation. Indeed, the amount of new combinations that sexual reproduction can produce - in other words, the amount of genetic variation that sexual reproduction can produce - is staggering.
So, a cell 2n = 4, has two pairs of homologus chromosomes. Let's call one pair 'A' and 'a' and the other pair 'B' and 'b'. ***Don't get confused by thinking that these letters represent single genes; they do not. Rather, they simply represent different chromosomes in a homologous pair. Because chromosomes carry 100's - 1000's of genes, it is almost impossible for homologs to be genetically identical over their entire length. So, although two homologs might carry the same gene at a particular locus (which we could, confusingly, represent as 'AA'), the homologs will NOT be the same across all the genes they carry. So, we represent their similarity as homologs by using the same letter, but their necessary DIFFERENCE with capital and lower case versions of that letter (Aa). OK?*** Now, you know that this organism can produce 4 gametes as a consequence of segregation and independent assortment: AB, Ab, aB, ab. A cell that is 2n = 6 has three pairs of homologs, AaBbCc. And, it can produce 8 types of gametes, ABC, abc, Abc, aBC, ABc, abC, AbC, aBc. So, there is a pattern here. The number of chromosomal combinations that are possible in the gametes = 2n (two to the 'n' power, where 'n' = the number of chromosomes in the haploid set). So, for 2n = 4, n = 2, and gamete number = 22 = 4. And, for 2n = 6, n = 3, and gamete number = 23 = 8. Well, most organisms have lots of chromosomes... humans have 2n = 46. So, n = 23, and the number of different types of genetic combinations that we EACH can make in our gametes is 223 = about 8 million. That's more than the population of Georgia or New Jersey. That is a staggering amount of genetic variation, produced by each single human being. But of course, one gamete does not make an offspring; you need two to tango (so to speak). So, if a male can produce 8 million different types of sperm, and if a female can produce 8 million different types of eggs, and if any of the sperm are just as likely to fertilize any of the eggs, then there are 8 million x 8 million possible combinations of sperm and egg (that's one big Punnett Square!); that's 223 x 223 = 246: about 70 trillion different types of zygotes that are possible. That's a LOT of variability, just from one pair of reproducing humans. Now, the differences in these 8 million gametes are slight. A couple of alleles, here and there. And, we are still talking about a human genome, so the entire set of genes still codes for a human; a gene for every trait. So, we are all fundamentally very similar to one another, sharing 99.9% of our genes. The ways we differ are actually pretty subtle... the color of hair, the slope of the nose, the bend of a finger. That's probably why other species seem so homogeneous to us until we really sit down and try to recognize these little personal differences. That's the solution to Darwin's Dilemma about the source of variation.... hereditary particles don't blend together; they remain distinct and form new combinations from generation to generation, LARGELY AS A CONSEQUENCE OF INDEPENDENT ASSORTMENT, BUT ALSO AS A CONSEQUENCE OF CROSSING OVER, WHICH WE WILL DESCRIBE LATER. REALIZE THAT THE ARGUMENTS ABOVE ARE ONLY LOOKING AT THE EFFECTS OF INDEPENDENT ASSORTMENT. This huge amount of variation is not caused by the production of a new gene by mutation (although this does occur); this remarkable amount of variation is produced by recombination: the formation of new combinations of genes in gametes and offspring during sexual reproduction.
3. Model of Evolution circa 1905 (Rediscovery of Mendel's Laws)
Source of variation: Independent assortment of particulate hereditary particles during gamete formation.
Agents of change: NS.
While Mendel's contributions were seminal, his postulates and principles did not describe the full range of complexity that occurs in the patterns of heredity or the expression of genetic information in the phenotype. Although many genes exhibit simple patterns of complete dominance, even segregation, and independent assortment, many do not. In addition, the genes that are received by an organism are not the sole determinant of the phenotype - even of the characteristics that these genes affect. For the remainder of this unit, we will look at how Mendelian patterns are modified (or even violated).
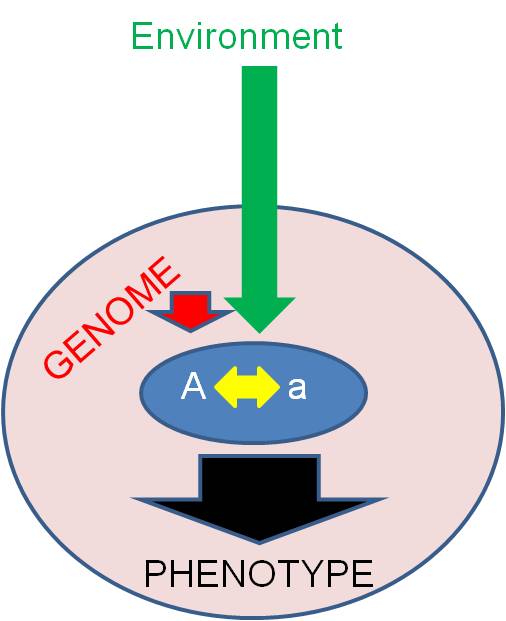 A.
Overview
A.
OverviewIn addition to considering whether (and how) an alelle is expressed, we will also consider the "value" of the allele to the organism (is it 'good' or 'bad'?) This may not be an intrinsic property of the allele. Rather, the value of an allele is also determined by these three classes of factors. Keep this in mind; it is the overriding message of transmission genetics. Unfortunately, our culture has not always appreciated these ideas.
Diploid organisms have two alleles for each locus, or gene. Intralocular effects describe the way that these alleles interact to cause a phenotype. You are familiar with one type of intralocular interaction already, so we will start there.
1. Complete Dominance (Mendelian)
In this case, the heterozygote expresses a phenotype indistinguishable from the phenotype of the homozygous 'dominant' individual.
- AA = Aa > aa (Mendel's peas provide examples) At a cellular level, why does this happen? Well, genes code for proteins. Apparently, one gene (A - the dominant gene) codes for the production of enough functional protein for complete cell function and phenotypic expression. Surplus has no effect. Maybe a reaction is limited by the amount of substrate, and not the concentration of enzyme (protein). So, one 'dose' of enzymes is enough to metabolize all of the substrate, and extra enzyme doesn't change this effect. Or the product of the dominant allele has a higher reactivity with the substrate and always reacts with it, it's way, even in the heterozygous condition.
 2.
Incomplete Dominance/Intermediate Inheritance In this case, the heterozygote expresses a phenotype that is intermediate between
the phenotypes expressed by the homozygotes. This is quite common. A classic
example is flower color in 4 o'clocks. Homozygotes for R ("RR") produce
red flowers, homozygotes for "r" (rr) produce white flowers, and heterozygotes
(Rr) produce pink flowers.
- AA > Aa > aa
At a cellular level, this is probably a function of a 'dosage effect'. Two
'on' genes produce more functional product than one, and this surplus influences
cell function and phenotypic expression. Maybe the protein product of an active
R gene is the pigment, itself, and r produces a nonfunctional protein (maybe
it has a premature stop codon and no protein is even produced). So, the two
functional genes in the RR homozygote produce more pigment and a deeper red
color than the single functional gene in the pink heterozygote (Aa).
2.
Incomplete Dominance/Intermediate Inheritance In this case, the heterozygote expresses a phenotype that is intermediate between
the phenotypes expressed by the homozygotes. This is quite common. A classic
example is flower color in 4 o'clocks. Homozygotes for R ("RR") produce
red flowers, homozygotes for "r" (rr) produce white flowers, and heterozygotes
(Rr) produce pink flowers.
- AA > Aa > aa
At a cellular level, this is probably a function of a 'dosage effect'. Two
'on' genes produce more functional product than one, and this surplus influences
cell function and phenotypic expression. Maybe the protein product of an active
R gene is the pigment, itself, and r produces a nonfunctional protein (maybe
it has a premature stop codon and no protein is even produced). So, the two
functional genes in the RR homozygote produce more pigment and a deeper red
color than the single functional gene in the pink heterozygote (Aa).
3. Codominance
In this case, the heterozygote expresses both traits completely; this does not produce something that is "in between" the homozygotes; rather, the heterozygote expresses both traits expressed in the homozygotes. The classic example is the A-B-O blood group in humans. These letters refer to alleles in the human population that encode a protein that is placed on the surface of blood cells. These proteins are called 'surface antigens', and they act as a chemical signal to white blood cells that these cells are "self". There are three alleles in the human population at this locus. Of course, each diploid person only has two of these three alleles. The genotypes that are possible, and the phenotypes they express, are as shown:
| Genotype | Interaction between alleles | Phenotype |
| AA | A | |
| AO | A dominant to O | A |
| BB | B | |
| BO | B dominant to O | B |
| OO | O | |
| AB | A and B codominant | AB |
The A allele codes for the A surface antigen. The B allele codes for the B
surface antigen. The O allele is non-functional; no surface antigens are produced.
So, any genotype with an A allele makes A surface antigen; any genotype with
a B makes B surface antigen. A genotype that is AB has different active genes,
each producing their own surface antigen, and so BOTH A and B surface antigens
are affixed to the outside of the cell. It is a phenotype that is BOTH A and
B at the same time.
A more general cellular explanation is this. The allelic products are both
functional and work on slightly different substrates. Having both gives the
phenotype a qualitative diffence, not a quantitative difference. This is often
confused with "incomplete dominance", but think about it this way:
"pink" flowers are neither "red" nor "white" -
they are something different that isn't red or white. But AB blood is BOTH A
and B at the same time, not different from A and B, but a combination of both
A and B. 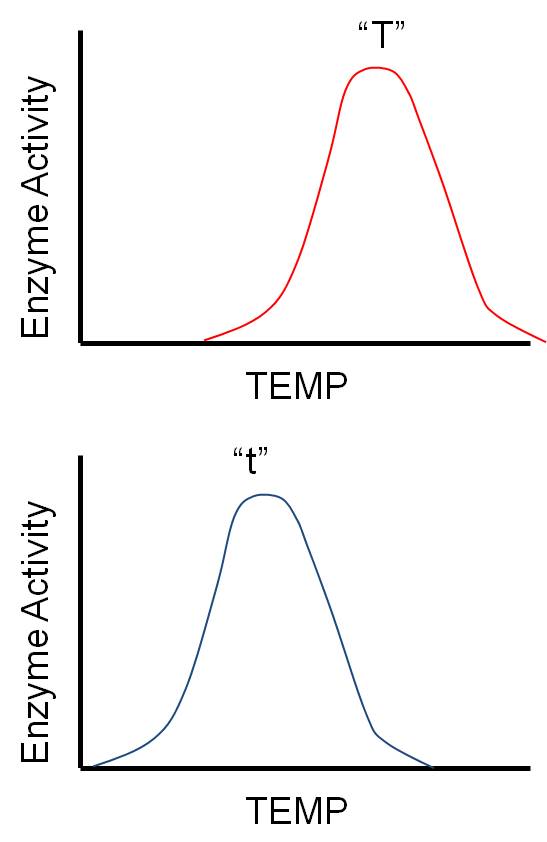
4. Overdominance (Heterosis) In this seemingly unusual situation, the heterozygote expresses a phenotype "more extreme" than either homozygote. So, if AA = tall and aa = short, then the heterozygote Aa = Tallest. This may seem unusual, but it is actually easy to explain. Suppose the two alleles work slightly differently, or under slightly different conditions. For instance, consider a gene that codes for an enzyme that influences growth. As you can imagine, there are LOTS of such genes! Almost any catalyst that affects metabolism, respiration or protein synthesis will influence growth. Maybe the two alleles for this enzyme work at different temperatures (H = warm and h = cold). If the organism lives in an environment that is warm most of the time, then the homozygote for HH will grow most of the time, and will be taller than the hh homozygotes that only grow during the rare colder periods. However, the heterozygote Hh grows ALL the time, when it is cold and warm, and so is taller than either homozygote. Variation at a locus can be as advantageous as variation in offspring; variation in the genotype (heterozygosity) may allow the organism to cope with a greater range of environmental conditions, or metabolize more substrates. Thus, there is often a 'heterozygote advantage' at certain loci.
As a consequence of many loci acting on a trait, there are many more combinations that are possible - resulting in a wide variety of phenotypic expression that forms a nearly 'continuous' range of variation.
For example, consider human skin pigmentation. There are at least 16 loci that code for melanin (skin pigment) production. Although there is complete dominance at each locus, this multiplication of genes allows for an extraordinary and continuous amount of phenotypic variation, from all 16 genes 'on' (and very very dark skin), to 15 'on' and 1 'off' ( very dark skin), to 14 'on' and 2 'off' (a bit lighter), to , 13:3, etc. , all the way to 0 'on' and 16 'off' (no pigments produced at all).
Humans evolved on the plains of Africa. The loss of hair was adaptive (to decrease insulation in the subtropical heat), but this made the skin vulnerable to UV rays that cause mutation. Selection favored humans that could make lots of melanin, to protect the skin against these UV rays. Selection favored humans that duplicated their melanin genes, and could thus produce more pigment. Humans that continued to live in tropical areas continued to benefit from this dark skin. In cooler climes with less intense sun, selection favored humans that spent less energy on melanin production, selecting for the recessive alleles at each of these multiple loci. In addition, lighter skinned people benefitted at these latitudes by using the sun's energy to synthesize vitamin D in the skin.
 In
epistasis, one locus has precedence over the expression at another locus and
can override it. Albinism is the classic example. Albinism ooccurs in all populations
of humans (and many other animals). It usually does NOT involve the melanin
producing genes. (So, true albinism is NOT caused by the 0 'on', 16 'off' scenario
described above). Rather, it involves another locus. An 'aa' individual at the
albinism locus does not make the precursor for melanin. So, with no
precursor from which melanin can be made, it doesn't matter WHAT the genotypes
are at the 16 melanin loci - there won't be any melanin produced. This is why
albinism occurs in all human populations (or 'races') regardless of of the skin
color typical for that population - because it is influenced by a gene that
is inherited independent of the melanin producing genes.
In
epistasis, one locus has precedence over the expression at another locus and
can override it. Albinism is the classic example. Albinism ooccurs in all populations
of humans (and many other animals). It usually does NOT involve the melanin
producing genes. (So, true albinism is NOT caused by the 0 'on', 16 'off' scenario
described above). Rather, it involves another locus. An 'aa' individual at the
albinism locus does not make the precursor for melanin. So, with no
precursor from which melanin can be made, it doesn't matter WHAT the genotypes
are at the 16 melanin loci - there won't be any melanin produced. This is why
albinism occurs in all human populations (or 'races') regardless of of the skin
color typical for that population - because it is influenced by a gene that
is inherited independent of the melanin producing genes.
Multiple loci may influence genes in interactive ways, not just in quantitative, additive ways like skin color. For example, in sweet peas (not the garne peas of Mendel), there are two genes that influence flower color. Both exhibit complete dominance, and both proteins must be produced in order for purple flower color to be expressed. So:
aaBB (white) x AAbb (white)
|
100% AaBb (purple)
|
9/16 A_B_ (purple)
7/16 aaB_ or A_bb or aabb (white) (In these examples, the underlined space means that it
doesn't matter what the second allele is... if one allele at a locus is dominant,
then in a completely dominant system, you will get the same phenotype regardless
of whether the second allele is dominant or recessive. So, rather than writing
all the separate genotypes out that yield the same phenotype, you just put the
genes that determine the phenotype. Of course, to express the recessive phenotype,
you have to be homozygous - so to represent genotypes that express the recessive
phenotype, you need to write both alleles. aaB_ refers to aaBB and aaBb... they
both yield the aB phenotype.)
 C.
Environmental Interactions
C.
Environmental InteractionsThe environment has many direct effects on the phenotype. A fox may change from white to brown because a bucket of white paint falls on it's head. That would be a direct environmental effect; it happened independently of anything going on with the genotype. Those are NOT the effects we are talking about here. Rather, we are talking here about an interactive effect between the environment and the genes at a locus, such that the environment changes how the genotype is expressed as the phenotype.
1. Temperature
In arctic fox, the brown summer fur turns to white as temperatures drop. This is NOT a direct effect of temperature change on pigment in the hair shaft. Rather, the change in temperature changes how the genes and their protein products work. As temperatures drop, melanin genes are turned off, or the enzymes that catalyze the production of melanin change shape and no longer function. The result is that no melanin is produced, and thus the fur turns white.
2. Toxins
Susceptibility to certain mutagens will vary with the genotype. So, two people who are homozygous for a type of lung cancer (and 'should' genetically express that cancer) might have very different phenotypes. One, who smokes and exposes their lungs to mutagenic compounds might trigger those cancer genes. The other person, however, with the same genotype, might not have cancer because they never smoked and thus never exposed themselves to the environmental "trigger". Likewise, someone homozygous for an alternative allele might smoke without developing lung cancer.
These effects may be "probabilistic". For instance, some genes associated with cancers are only expressed (cause cancer) in a fraction of the individuals with that genotype. This could be due to different environmental exposure, but it could also be caused by different genes at other loci that interact with this cancer gene and augment or supress its effects through interlocular interactions.
D. The 'Value' of an Allele
Environmental Effects: The value of an allele often depends on the environment
In Sickle Cell Anemia, red blood cells 'sickle' when the concentration of oxygen in the bloodstream drops. This occurs when the person is active and the oxygen demand by muscle increases, drawing oxygen out of the bloodstream and depressing oxygen concentration in the cells in the bloodstream. The sickle-shaped cells do not pass through capillary beds as easily as normal red blood cells; they clog capillaries, resulting in oxygen deprivation and tissue damage downstream from the clot. This is usually most pronounced in the liver, kidneys, and brain, and eventually often results in premature death.
Sickle cell anemia is caused by an altered beta globin allele, which causes a single amino acid change in the beta-globin proteins in hemoglobin. The trait exhibits incomplete dominance - one "s" allele will result in some "sickling" of red blood cells at low oxygen concentration, but the condition is not nearly as severe as it is in the homozygous condition.
Now, you might think that even one sickle-cell allele would 'always be bad' - after all, I just said that heterozygotes do suffer some debilitating effects. But in this case, the "value" of a sickle cell allele - whether it is 'good' or 'bad' - depends on the environment. In particular, it depends on the presence of the Plasmodium parasite that causes malaria.
In the tropics, a primary source of human mortality is malaria. In 2006, nearly 900,000 people died of malaria; over 91% of these deaths were children in Africa. There were an estimated 250 million cases reported globally. Malaria is caused by several species of protists in the genus Plasmodium. This single-celled parasite is transmitted by female mosquitos. When they bite a human to take a blood meal (only female mosquitos drink blood - they use the protein to nourish their developing eggs), the parasites enter the human host's bloodstream and infects red blood cells. They divide mitotically, producing hundreds of offspring and eventually rupturing the cell. Thus, infection causes extreme loss of RBC's - "anemia" (don't get this confused with sickle cell anemia; we are just talking about malaria right now!!). Curiously, the Plasmodium parasite can not reproduce in cells with the altered form of "sickle-cell" hemoglobin - even in the heterozygous condition. So, the heterozygote suffers some sickling on occasion, but is protected from malaria. In Africa, SS homozygotes have lower survivorship than the Ssheterozygotes because the SS individuals are exposed to malaria. The ss homozygotes have lower survivorship than the Ss heterozygotes because of the more pronounced debilitating effects of the sickle cell disease.
So, is an 's' allele "good" or "bad"? Well, that depends on other alleles at that locus (if it's with another 's' allele it is always bad), but it also depends on the environment. If the 's' is with an 'S' in the temperate zone, it is bad (relative to the reproductive succes of the normal SS homozygote).... but if the 's' allele is paired with an 'S' allele in the tropics, then it is "good" - better than having two dominant alleles (SS).
TEMPERATE ZONE: survivorship: SS > Ss > ss
TROPICS: survivorship: SS < Ss > ss
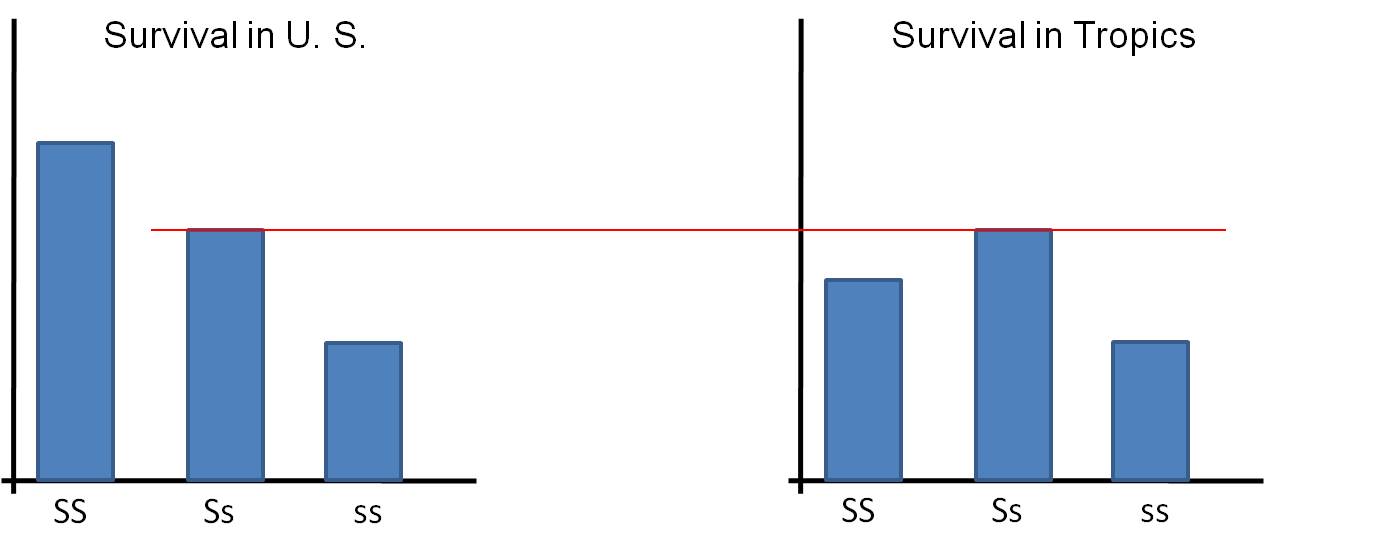
You should relate this to the corollary of Darwin's Theory of Natural Selection.
Populations in different environments will diverge from one another genetically,
as the environment selects for different traits (genotypes).
E. Summary:
An organism is more than just the sum of their parts, even at a genetic level. How the genotype works is not just the additive sum of all the genes acting independently. Rather, the way some genes work depends on the other genes in the genotype. The phenotype - what a complex organism IS - is the result of these complex interactions between genes, with the additional layer of environmental interactions and direct environmental effects. Indeed, given the potential complexities involved in how 1000's of genes and a complex environment can interact, it is rather surprizing that many genetic effects are simple enough to model as independent entities with a Punnet Square - without considering the other genes or the environment. So, Mendelian Genetics are actually the easiest, simplest patterns in heredity to recognize: and that is probably why they were recognized first.
Study Questions:
1. Understand Mendel's four postulates. State his 'first principle' of heredity without using the word 'segregate' in your definition, and his 'second principle' without using the words 'independent assortment' in your definition.
2. Know why Mendel conduct reciprocal crosses. What hypotheses did this test?
3. Understand how the results of his dihybrid cross lead him to hypothesize that the traits must be inherited independently. Understand the product rule.
5. Consider this cross: (male) AaBbCcDD x (female) AaBBccDD
- Assume independent assortment and dominance (capital is dominant to lower case) at each gene.
- How many types of gametes can each parent make with respect to these traits?
- How many genotypes and phenotypes are possible in the progeny?
- What fraction of offspring would you expect to express the ABCD phenotype?
6. Consider this cross:
AaBbCc x AaBbCC
- assume independent assortment of the three genes
- There is incomplete dominance at the A locus (meaning A is incompletely
dominant to a).
- There is complete dominance at the B locus.
- There is overdominance at the C locus.
How many genotypes are possible in the offspring?
How many phenotypes are possible in the offspring?
7. Provide a cellular explanation for codominance and overdominance.
8. Know how to diagram a cell, 2n=4, as it enters and leaves each cycle of Meiosis.
9. Explain how the movement of chromosomes in Meiosis I provides a mechanistic description of how Mendel's two principles work.
10) know they different types of allelic interactions, and WHY they occur, at a cellular (enzymatic) level.
11) Know the difference between quantitative traits and epistatic effects - and be able to explain the difference at a cellular (enzymatic) level.
12) Know how the environment can influence the expression of a gene; again, at the enzymatic level.
13) Know how the environment can influence the VALUE of a trait? Relate this to Darwin's idea of the diverge of populations in different environments.