 A.
Mendel's Life (from Villanova
University)
A.
Mendel's Life (from Villanova
University)
In this unit, we will examine how DNA is transferred through time, we will describe the process of sexual reproduction, and we will solve Darwin's Dilemma regarding the source of heritable variation. In addition, we will examine how the activity of genes is regulated - a process that results in the coordinated specialization of cell types and tissues through time (development).
A.
Mendel's Life (from Villanova
University) Johann Mendel was born in Moravia (now the Czech Republic) on July 22, 1822, into a poor farming family. He received an excellent education at the local schools, and entered the Augustinian order at the St. Thomas monastery in Brunn (now Brno) in1843. He was ordained in 1847 and was given the name 'Gregor'. The monastery sent him the the University of Vienna for two years (1851-1853), where he trained to be a teacher in physics and mathematics. After returning to Brunn, he began his experiments and taught part-time. He presented his work at scientific meetings in 1865, and the work was published in the proceedings of the meetings in 1866. His paper, Versuche über Pflanzen-Hybriden ("Experiments in Plant Hybridization,") provided one othe first quantitative examinations of a biological phenomena. The conclusions and implications of the work went largely unnoticed for 30 years, until the work was cited by three botanists independently working on plant hybridization in 1900. Mendel became the Abbot of the monastery, and his administrative duties put an end to his career as an experimental biologist. Mendel died in Brünn on 6 January 1884. Before his death, he stated, "My scientific labors have brought me a great deal of satisfaction, and I am convinced that before long the entire world will praise the result of these labors." His conviction came to pass, as he is now recognized as the founding father of genetics.
It was common knowledge that offspring looked alot like their parents. This observation was especially obvious and important in the agrarian societies of the pre-industrial age, where people saw that certain traits in domesticated plants, domesticated animals, and humans 'ran in families'. In fact, this was not just an idle issue. Understanding how heredity worked was an important economic question to these farmers, who were trying to breed more valuable and productive crops and livestock.
1. Preformationist Ideas
 One
school of thought - the "preformationist" school, thought that a miniature
human lived within either the egg (the 'ovist' school), or the sperm ('the homunculan'
school). This offspring laid in a state of suspended animation until the sperm
stimulated the egg to grow (for the ovist school), or the sperm was placed in
the fertile womb to grow ('homunculan' school). Of course, both of these ideas
had serious flaws. If the offspring was really just a product of one sex, then
why did the offspring have characteristics of both parents? Some theologians
saw interesting implications, too. If all generations were preformed, then each
generation must be nested within the egg or sperm of the preceeding generation
- also already preformed. The generations of humans would be like a set of russian
dolls, one nested with the other. At some point, it would seem that nature would
reach the limit of how small a preformed person could be - and that would be
the last generation (which would be followed by the apocalypse).
One
school of thought - the "preformationist" school, thought that a miniature
human lived within either the egg (the 'ovist' school), or the sperm ('the homunculan'
school). This offspring laid in a state of suspended animation until the sperm
stimulated the egg to grow (for the ovist school), or the sperm was placed in
the fertile womb to grow ('homunculan' school). Of course, both of these ideas
had serious flaws. If the offspring was really just a product of one sex, then
why did the offspring have characteristics of both parents? Some theologians
saw interesting implications, too. If all generations were preformed, then each
generation must be nested within the egg or sperm of the preceeding generation
- also already preformed. The generations of humans would be like a set of russian
dolls, one nested with the other. At some point, it would seem that nature would
reach the limit of how small a preformed person could be - and that would be
the last generation (which would be followed by the apocalypse).
2. Epigenetic Ideas
The other major school of thought promoted the "epigenetic" idea. Here, the egg does not actually contain a small individual, but rather only the potential to grow and develop into an individual. This idea had much less support, because there were no ideas about how this could happen; the growth of a small human was alot easier to understand than the development of organs and organ systems from 'nothing' - or at least, from things that weren't organs or organ systems! However, the epigenetic idea had one major benefit - it could explain why offspring often expressed characteristics of both parents. If both parents contributed 'stuff' to this epigenetic mass of potential, then the developing offspring could inherit traits from both parents. Because semen is a fluid, it seemed likely that the genetic information was a fluid. The 'mixing' of the heredity fluids from two parents might explain, sort of, why the offspring expressed a mixture of traits from both parents.
There was an additional observation that was very problematic with this blending idea, however. Most people were well aware that some traits 'skipped a generation', or even many generations. The reappearance of an ancestral trait (" you have your great-grandmother's nose") was know as a 'sport'. If the hereditary information from two parents were fluids that mixed, like black and white paint, it seemed difficult to explain how, after mixing black and white into grey, you could get some pure white paint back out of the mixture in a later generation.
 Mendel
set out to test these alternate hypotheses by breeding organisms from different
'pure-breeding' stocks together. In other words, Mendel conducted hybridization
experiments, forming hybrids by crossing strains that bred true for particular
characteristics. He selected the common garden pea, Pisum sativum,
as his study organism. He chose this plant because there were alot of pure-breeding
varieties already available; farmers had selected for different characteristics,
creating strains that produced round peas, wrinkled peas, yellow peas, green
peas, tall plants, short plants, etc. Indeed, Mendel reports that he received
34 different strains from farmers. In addition, because peas were fast growing
and fecund (produced lots of offspring - each pea is a separate offspring),
he could generate a lot of data in a matter of months. (Some characters, however,
like flower color or plant height, could only be determined in the following
year, when the offspring grew up and exhibited the trait.) Being trained in
mathematics and probability, he knew that a large sample would provide a 'truer'
indication of a pattern than a small sample.
Modern geneticists have chosen other study organisms - like fruit flies, mice,
and bacteria - for the same reason.
Mendel
set out to test these alternate hypotheses by breeding organisms from different
'pure-breeding' stocks together. In other words, Mendel conducted hybridization
experiments, forming hybrids by crossing strains that bred true for particular
characteristics. He selected the common garden pea, Pisum sativum,
as his study organism. He chose this plant because there were alot of pure-breeding
varieties already available; farmers had selected for different characteristics,
creating strains that produced round peas, wrinkled peas, yellow peas, green
peas, tall plants, short plants, etc. Indeed, Mendel reports that he received
34 different strains from farmers. In addition, because peas were fast growing
and fecund (produced lots of offspring - each pea is a separate offspring),
he could generate a lot of data in a matter of months. (Some characters, however,
like flower color or plant height, could only be determined in the following
year, when the offspring grew up and exhibited the trait.) Being trained in
mathematics and probability, he knew that a large sample would provide a 'truer'
indication of a pattern than a small sample.
Modern geneticists have chosen other study organisms - like fruit flies, mice,
and bacteria - for the same reason.
1. Monohybrid Experiments (single traits)
Mendel reported the results of seven parallel experiments. In each experiment, he cross plants that had different expressions of a particular trait. So, in the third set of trials, he bred plants that 'bred true' for white flowers (when bred with themselves) with plants that produced purple flowers. For this experiment, he did 35 cross-fertilizations with ten plants.
a. Reciprocal parental crosses tested the ovist and homunculan schools:
A very important methodological aspect of the experiment was that he conducted reciprocal crosses between stocks. So, he placed pollen pure-breeding white flowers on the stigma of purple-flowers, and pollen from purple flowers on the stigmas of white flowers. So, by comparing the results of these reciprocal crosses, he could see if the inheritance of the trait (flower color) was associated with the sex of the parent. If the homunculan school was correct, then the offspring should bear the trait expressed in the stock that donated the pollen. If the ovist school was correct, then the offspring should bear flowers the color of the strain that received the pollen and donated the egg (in the ovule).
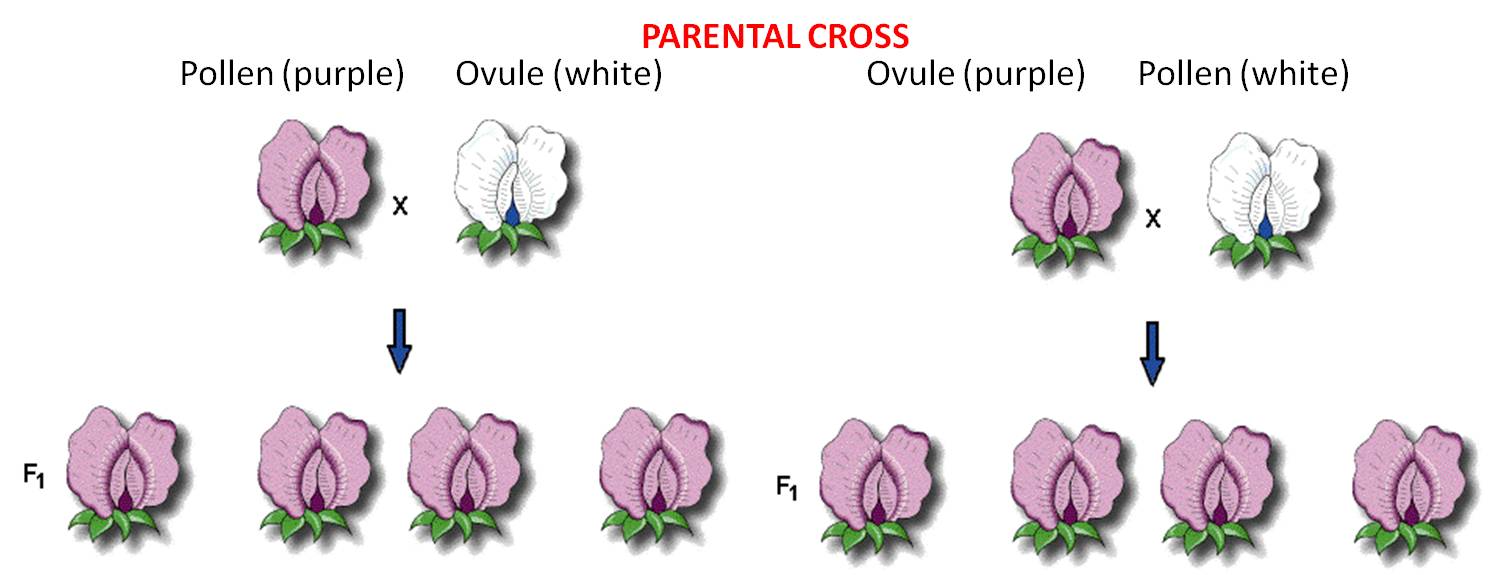 These 'pure-breeding'
stocks were called the 'parental generation'.
These 'pure-breeding'
stocks were called the 'parental generation'.
pollen (male) from Purple flowers X ovule (female) from White flowers
AND
pollen (male) from White flowers X ovule (female) of Purple flowers
These crosses produced offspring (peas). The first generation of 'hybrids' produced from pure-breeding parental strains is called the F1 (for 'first 'filial') generation. He planted and grew the peas produced from both crosses and observed their flower color. In both reciprocal crosses, all the plants produced purple flowers. These results falsified both the homunculan and ovist schools; both hypotheses predicted that plants from one or the other cross would yield white flowers. Indeed, Mendel realized that these results suggested that both parents had to contribute hereditary factors for this trait; the purple offspring from the first reciprocal demonstrated that the male contributes, and the purple offspring in the other reciprocal demonstrated that the female contributes, too. But if both parents contributed hereditary information, what happened to the white information contributed by the other parent in each cross? Mendel hypothesized that the contribution was present in the offspring, but not expressed. He coined the term 'dominant' for the trait expressed in the F1, and 'recessive' for the trait he thought was there, but hidden. He became more confident in these hypotheses when all seven traits that he studied showed the same pattern.
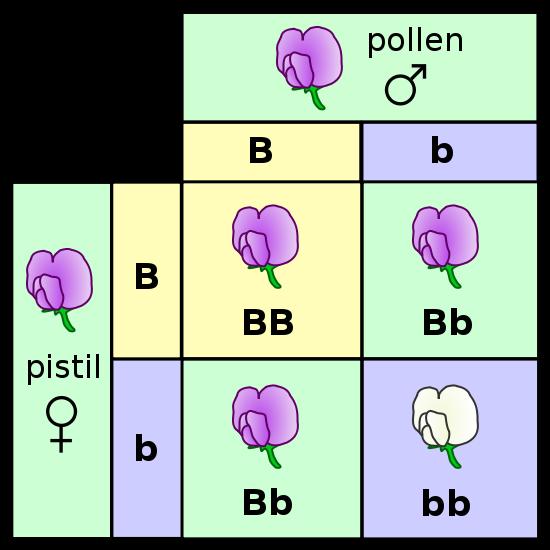 b.
Crossing the F1 hybrids tested the blending hypothesis:
b.
Crossing the F1 hybrids tested the blending hypothesis:
He allowed the F1 plants to fertilize themselves, essentially performing an F1 x F1 cross. He makes reference to the fact that, in many species and for many traits, the hybrids (F1's) exhibit a triat intermediate to the two parental types. This intermediate result might be expected of a blending pattern (white paint and black paint making grey), but one trait dominating the other could also be produced from a blending mechanism, too. One thing you would not expect from a blending of fluids, however, would be the re-expression of the recessive 'white' trait. Grey paint mixed with grey paint should not produce white paint.
The peas from these crosses were grown and the flowers were observed. Of the 929 plants that he grew from these seeds, 705 bore violet flowers and 224 had white flowers; a ratio of 3.15:1. In fact, all seven traits produced ratios near a 3:1 ratio in the F2 generation (offspring of F1 x F1 crosses). He recognized the importance of the 3:1 ratio when combining two things. It reminded him of a simple binomial expansion (a + b)2 = 1aa + 2ab + 1bb. If an organism has 2 different types of hereditary units (a = purple and b = white) for a given trait and it mates with itself, then there are three combinations that are possible in the offspring (aa, ab, bb). And, they should occur in a 1/4:2/4:1/4 ratio. If, as a consequence of dominance, the progeny with different particles (ab) only express one - the 'a' (purple) - then they will look like the aa's and 'a' (purple) offspring will occur 3/4 of the time and 'b' (white) offspring will occur only 1/4 of the time.
c. Mendel proposed four postulates (hypotheses) to explain these data:
- genetic info is unitary or 'particulate' - it is not a fluid that blends
- each oganism has two unit factors (we call these 'genes' now) for each trait
- if an organism has different factors for a given trait, one is expressed (dominant) over the other (recessive)
- the two genes for a given trait separate and go into separate gametes during gamete formation. Subsequent fertilization (fusion of gametes) is random. This is called Mendels First Principle: Principle of Segregation
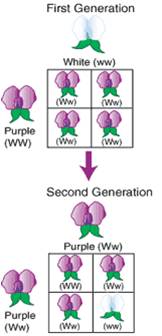 d.
How his hypothesis explains his observations:
d.
How his hypothesis explains his observations:
Parentals: WW (Purple) x ww (white) .... (and the reciprocal)
Gametes: W w
F1 offspring:
Genotypic Ratio: 100% Ww (heterozygous)
Phenotypic Ratio: 100% Purple
F1 x F1 Cross: Ww x Ww
Gametes:
W and w W
and w
F2 offspring:
Genotypic Ratio: 1/4 WW, 2/4 Ww, 1/4 ww Some Terms:
Phenotypic Ratio: 3/4 Purple, 1/4 White
Genotype = the type of genes an organism has, like Ww.
Genotypic Ratio = is the fractional representation of different
genotypes in a group, such as 1/4WW : 1/2Ww : 1/4 ww
Phenotype = the characteristic or trait that is expressed,
such as 'Purple'. Usually, this is represented by the gene that causes the trait,
such as 'W' in the example, above.
Phenotypic Ratio = the fractional representation of different
phentypes in a group, such as 3/4W : 1/4 w.
2. Monohybrid Test Crosses
If the hypotheses are correct, Mendel reasoned, then the F1 has a gene for the recessive 'white' trait that is not expressed. How could he determine whether it was there or not? He realized that he could answer this question if he crossed it to the recessive white parent: (ww). The logic is this: the homozygous recessive parent can only pass recessive alleles to the offspring. Whatever the offspring receives from the OTHER parent will determine the expression of the trait. It the offspring receives a dominant allele for purple from the other parent, then the offspring (Ww) will express purple flowers. But, if the other parent is hiding a recessive allele and passes that to the offspring, then the offspring will have the homozygous recessive genotype (ww) and will produce white flowers.
Mendel predicted that, under this hypothesis, the phenotypic ratio in the offspring would be: 50% purple (W): 50% white (w). The results supported his hypothesis, and supported his postulates. Replication of this results across all seven traits gave strong confirmation that he had described a general pattern of heredity.
More
Terms:
Homozygous: A zygote is a fertilized egg that has received
genes from both parents. If the genes received from both parents for a given
trait are the same, then this zygote (and the organism that develops from it)
is homozygous for this trait. If the zygote has received dominant genes
for the trait from both parents, it is homozygous dominant (WW); if
it has received the recessive allele from both parents, it is homozygous
recessive (ww).
Heterozygous: If the zygote receives different alleles from
the parents for a given trait, then the zygote is heterozygous for
that trait (Ww).
Alleles are different 'forms' of a gene, that affect a given
trait - like flower color - in different ways ('W'and 'w' are alleles for flower
color).
3. Dihybrid Experiments
Most animal and plant breeders had noticed that some traits seem to be inherited together. Indeed, in his own experiments, Mendel saw that flower color was always correlated with the color of the seed coat, such that seeds with white seed coats grew into plants with white flowers, and seed with brown seed coats grew into plants with purple flowers. Having demonstrated that his seven traits all exhibited the same pattern of inheritance, he now set out to determine whether these traits were inherited in a dependent or independent manner. This is a very insightful question. For example, he had already demonstrated that yellow pea color was dominant to green, that round pea shape was dominant to wrinkled, and that F1xF1 crosses for each trait produced a 3:1 ratio in the offspring. Well, maybe these traits are inherited in the same way because they are governed by the SAME heritable unit factor (gene)... and that's why they produce the same pattern. If yellow and round were caused by the same gene, then of course they would be inherited in the same way. Or, maybe they are governed by different unit factors that have a pattern of heredity that is related to one another... maybe they are governed by different factors that "travel" from parent to offspring together - yellow always traveling with round, for instance. This had important practical applications, too. A sheep farmer would profit from sheep that produce both good wool and good milk (for feta or roquefort cheese) from his sheep herd. But, maybe the trait of quality wool is inherited with the trait of poor milk production. In that case, they might not want to strongly select for great wool quality, as milk production would suffer. It might be best to maintain a variable population that produces good wool and adequate milk. Every organism expresses a combination of traits, so addressing how combinations of genes are inherited is a very important question.
a. Parental crosses of stocks breeding true for two traits
First, Mendel bred plants together and created stocks that bred true for two traits: seed color and seed shape. He created a stock of plants that produced yellow, round peas and a stock that produced green, wrinkled peas. He already knew the dominance patterns, and had already concluded that parents bore two 'unit factors' (genes) for each trait. So, he hypothesized that the genotypes of the plants in each pure-breeding stock would be RRYY (round, yellow) and rryy (wrinkled green). He placed pollen from the wrinkled, green stock on flowers of the yellow, round stock and observed the progeny.
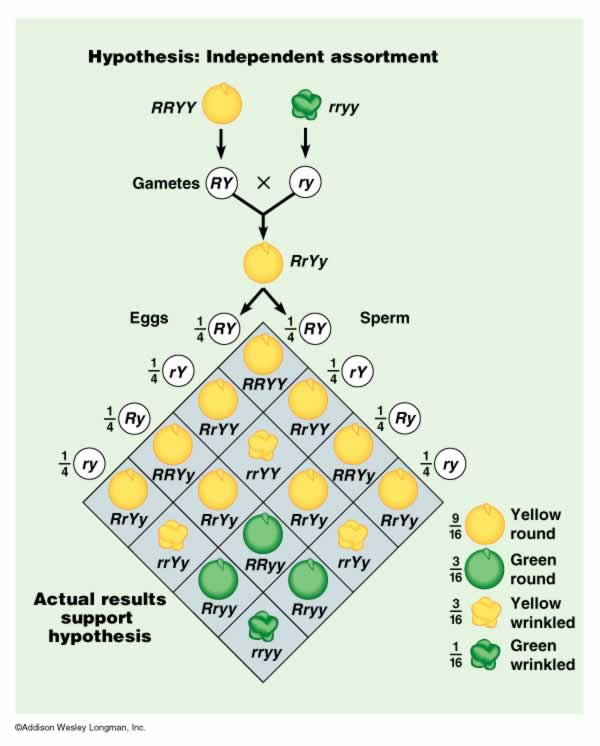 Round, Yellow (RRYY) x wrinkled, green (rryy)
Round, Yellow (RRYY) x wrinkled, green (rryy)
He hypothesized that the parental types would produce RY gametes and ry gametes, respectively. The combining of the genes in the offspring would make only RrYy genotypes, that should all express the phenotypic traits of yellow, round peas. This hypothesis was confirmed. However, this did not yet address the issue of independence; this pattern would also have occurred if color and shape were governed by one gene, or if the genes were inherited in a dependent manner. In short, the Y had to be inherited with an R, because that's all the RRYY parent had to give.
b. Crossing the F1's:
He crossed the F1 individuals, and found the following results:
- 315 round, yellow peas (~ 9/16)
- 101 wrinkled yellow peas (~3/16)
- 108 round green peas (~3/16)
- 32 wrinkled green peas (~1/16)
TOTAL - 556 peas
c. His interpretation and hypothesis:
First, it was obvious that the two traits were not caused by the same gene; all yellow peas were not ALSO round - he had produced some yellow wrinkled peas. So, different genes governed see color and seed shape, and so the genes for these traits could occur in differnt combinations.
Mendel realized that the 3:1 ratios were still preserved for each trait, when considered separately. For example, taken together, the ratio of yellow:green peas was 416:140 (2.97:1) and the ratio of round:wrinkled peas was 423:133 (3.18:1).
And also, Mendel realized that the
results of this experiment, and the results of subsequently self-crossing these
hybrids, were consistent with the probability of occurrence of independent events.
We know this as The Product Rule: "The probability that independent events
will occur together is the product of their independent frequencies."
These ratios in which the traits occurred in combination equaled the products of the independent frequencies:
3/4 W X 3/4 Y = 9/16 WY (round yellow)
1/4 w X 3/4 Y = 3/16 wY (wrinkled yellow)
3/4 W x 1/4 y = 3/16 Wy (round green)
1/4 w x 1/4 y = 1/16 wy (wrinkled green)
Mendel's hypothesis can be stated like this:
During gamete formation, the way one pair of genes (governing one trait) segregates is not affected by (is independent of) the pattern of segregation of other genes; subsequent fertilization is random. This is called Mendel's Second Principle: Principle of Independent Assortment.
He tested this hypothesis is subsequent experiments using three traits (seed color, seed shape, and seed coat color), and showed that the combination of traits expressed in the offspring were consistent with predicitons from their independent frequencies. For example, In a "trihybrid" F1 cross (a self-cross of an individual that is heterozygous for all three traits - YyRrCc), the fraction of peas that are yellow, round, and have a brown seed coat (caused by the dominant allele, "C") should be 3/4 x 3/4 x 3/4 = 27/64. His results confirmed his hypothesis.
4. Dihybrid test cross:
We can test for independence a bit more directly, through a dihybrid test cross. The principle of independent assortment is really predicting what the frequencies of different gamete types will be. So, consider an organism that is a double heterozygote (RrYy). The principle of independent assortment predicts that the segregation of the R genes (R and r) during gamete formation will be unrelated to (independent of) the pattern of segregation of the Y genes (Y and y) going into the same gametes.
With respect to the "R" gene, it has the Rr genotype and should produce R and r gametes at a ratio of 1/2 R to 1/2 r.
Likewise, with respect to the "Y" gene, the organism should produce Y and y gametes at a ratio of 1/2 Y to 1/2 y.
Now, every gamete must carry one (and only one) gene for each trait. So first of all, there can't be Rr or Yy gametes. And, under the hypothesis of independent assortment, the types and frequencies of the gene combinations that occur in the gametes can be predicted as the product of their independent frequencies. So, we would predict that this organism would produce 1/4 RY gametes (1/2 R x 1/2 Y), 1/4 Ry gametes (1/2 R x 1/2 y), 1/4 rY gametes (1/2 r x 1/2 Y), and 1/4 ry gametes (1/2 r x 1/2 y). These gametes are in the first column in the figure, below.
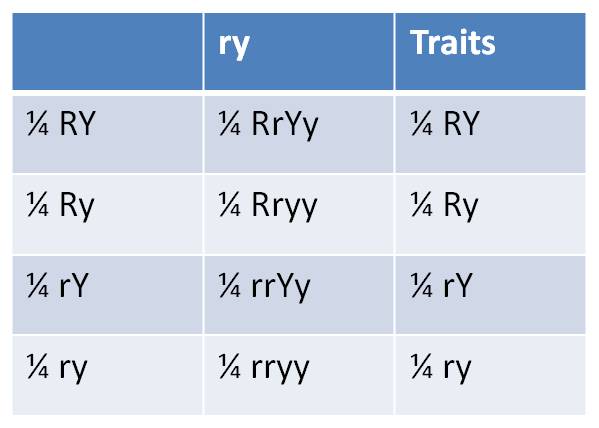 So,
to test independent assortment, we really want to know whether this organism
is producing gametes in these predicted frequencies. But we can't see the genes
in the gametes too easily; through heredity, all we can observe are the traits
in the phenotype of the offspring. And since these gametes carry recessive alleles,
it seems likely that they would be hidden from our observations.
So,
to test independent assortment, we really want to know whether this organism
is producing gametes in these predicted frequencies. But we can't see the genes
in the gametes too easily; through heredity, all we can observe are the traits
in the phenotype of the offspring. And since these gametes carry recessive alleles,
it seems likely that they would be hidden from our observations.
Unless we are clever.
If we mate this first organism to a second organism homozygous for the recessive traits (rryy), then the second parent will only donate recessive alleles (ry) to the offspring. (This gamete is in the header of the second column, at right). In this "test cross", the genes the offspring receives from the first parent will be expressed in the phenotype - we will be able to "see" the gene combinations, and their frequencies, produced in the gametes of the first individual. (The 'traits' expressed by the offspring - shown in the third column - are the same traits in the same frequencies as the gamete types produced by the first parent - shown in the first column.)
Mendel's quantitative approach and background in probability allowed him to conduct and interpret hybridization experiments in a new, more rigorous way. By examining and classifying the characteristics in offspring from particular parents, he was able to make insights into processes occuring at the cellular level. His two principles of heredity describe the allocation of genes to gametes - a process he never observed. However, 30 years later, cellular biologists confirmed that the hereditary units - called chromosomes - were allocated to gametes in a pattern consistent with Mendel's conclusions. It is worth reflecting on this for a second. All Mendel did was mate plants, collect and count seeds, and plant them. Yet, through his clever experimental design, he was able to test hypotheses and reach conclusions about phenomena that he could not observe directly. Human genius can be applied to any endeavor. In our modern technological society, we seem most impressed when genius is applied to making new technological innovations and devices. In science, however, genius can be expressed in the creativity of designing the correct experiment.
1. If the genes assort independently, then you can calculate ‘single gene’ outcomes and multiply results together…
For Example: AaBb x Aabb:
- what is the probability of an Aabb offspring?
- What is the probability of an offspring expressing Ab?
- How many genotypes are possible in the offspring?
- how many phenotypes are possible in the offspring?
Do the single-gene Punnett squares:
 and
and 
Answer each question for each gene, and then multiply the answers together:
- what is the probability of an Aabb
offspring? 1/2 (Aa) x 1/2 (bb) = 1/4
- What is the probability of an offspring expressing Ab? 3/4
(A) x 1/2 (b) = 3/8
- How many genotypes are possible in the offspring? 3
x 2 = 6
- how many phenotypes are possible in the offspring? 2
x 2 = 4
2. You can easily address more difficult multigene problems:
For Example: (female) AaBbCcdd x AABbccDD (male)
- how many types of gametes can each
parent produce?
- What is the probability of an offspring expressing ABCD?
- How many genotypes are possible in the offspring?
- how many phenotypes are possible in the offspring?
Answer the questions for each gene, then multiply the answers together:
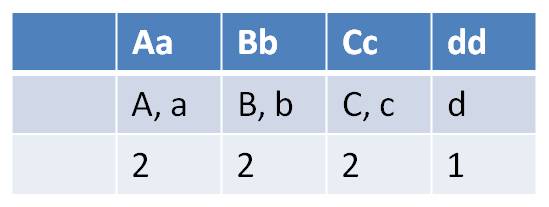
- how many types of gametes can each parent produce?
| FEMALES - types of gametes at each gene (locus) | MALES - types of gametes at each gene (locus) |
 |
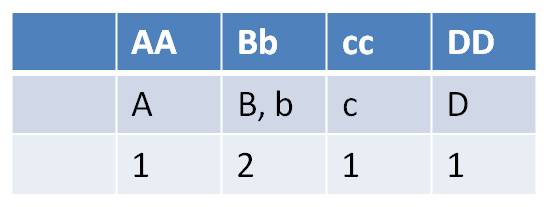 |
| Females can make 2 x 2 x 2 x 1 = 8 gamete types | Males can make 1 x 2 x 1 x 1 = 2 gametes types |
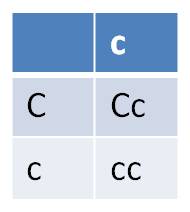
With respect to offspring, do the single-gene Punnett squares:
 |
 |
 |
 |
- What is the probability of an offspring
expressing ABCD? = 1 (A) x 3/4 (B) x 1/2 (C) x
1 (D) = 3/8
- How many genotypes are possible in the offspring? =
3 x 3 x 2 x 1 = 18
- how many phenotypes are possible in the offspring? =
2 x 2 x 1 x 1 = 4
Note that for the C and D genes, we simplified the Punnett Squares. If a parent is homozygous for a gene, then it can only make one type of gamete with respect to that trait. So, to reduce redundancy, we just used one column for the 'cc' parent in the third Punnett Square, and only one row and one column for the parents in the last Punnett Square. The same could have been done for the AA parent in the first Punnett square - the information in the two columns is identical.
Study Questions:
1. List Mendel's four postulates, clearly and specifically stating his 'first principle' of heredity without using the word 'segregate' in your definition..
2. Why is a test cross an effective way of determining the genotype of an individual?
3. Determine the genotypic and phenotypic ratios from the following crosses. Assume dominance of capital letter:
a. AA x AA
b. AA x Aa
c. Aa x Aa
d. Aa x aa
e. aa x aa
4. Why did Mendel conduct reciprocal crosses? What hypotheses did this test?
5. Why did Mendel conduct an F1 x F1 cross (the second generation of crosses)? What hypothesis did this test?
6. How did the results from the reciprocal crosses suggest that both parents must contribute genetic information?
7. What question was Mendel addressing with his dihybrid cross?
8. How did the results lead him to hypothesize that the traits must be inherited independently? Use the product rule in your answer.
9. State Mendel's second principle, without using the word 'independent' in your definition.
10. Consider this cross: (male) AaBbCcDD x (female) AaBBccDD
- Assume independent assortment and dominance (capital is dominant to lower case) at each gene.
- How many types of gametes can each parent make with respect to these traits?
- How many genotypes and phenotypes are possible in the progeny?
- What fraction of offspring would you expect to express the ABCD phenotype?