

The modern synthesis and population genetics, following on classical Mendelian genetics, tended to focus on evolution as a process that occurred as a consequence of changes in the gene frequencies of structural genes - genes that code for enzymes and structural proteins. And although these protein sequences do vary over evolutionary time, and although many changes in these genes are adaptive responses to the environment, the large scale changes in evolution - like the evolution of legs in the first amphibians or wings in the first birds - seemed to be due to another cause. Why? Because fish and birds are made out of the same proteins as their ancestors (fish and archosaurs, respectively). Subtle changes in the type of collagen protein produced, or the type of hemoglobin produced, could not explain these large scale changes in morphology that are truly the most dramatic patterns in evolution. Evolving a leg from a bony fin is not a function of making a slightly different type of collagen - it is a function of putting the collagen and cells together in a new way, to create a new shape. Indeed, some biologists were so convinced that these processes were fundamentally different that they use the term 'microevolution' to explain differences within a population and 'macroevolution' to explain large scale morphological changes over geologic time.
|
|
|
| Can changes in the frequency of genes that code for different proteins within populations, even if they are adaptive like flower color, explain major differences in body plan? | The structural proteins that make up these organisms are largely the same. The differences seem to be more a function of the way the proteins and cells are put together: the way the organism develops and expresses its morphology. | It turns out that these developmental plans are coordinated by genes, too. So small changes in developmental genes can have large effects on the final shape and characteristics of the organism. |
Ever since the 1800's, embryologists
were impressed by the pattern of epigenetic development - the formation of structures
by embryos that did not have them in the past. So, how does a frog embryo (like
the one below) that does NOT have legs today grow them tomorrow? During the
20th century, some evolutionary biologists suggested that the answer to this
question might have a bearing on how legs evolved in the first place...  how
did fish, which do NOT have legs, evolve into amphibians that DO have legs?
Starting in the 1970's, developmental biologists began to identify the genes
responsible for these changes in the 'shape' of organisms - genes that influenced
how balls of cells (embryos) developed into fish or amphibians, reptiles or
birds. And sure enough, the
insights gained in how an individual embryo grows legs gave insights into how
legs may have evolved. Studying the relationship between evolution and the genes
that regulate development became a field in itself: evolutionary development
or "evo-devo", for short.
how
did fish, which do NOT have legs, evolve into amphibians that DO have legs?
Starting in the 1970's, developmental biologists began to identify the genes
responsible for these changes in the 'shape' of organisms - genes that influenced
how balls of cells (embryos) developed into fish or amphibians, reptiles or
birds. And sure enough, the
insights gained in how an individual embryo grows legs gave insights into how
legs may have evolved. Studying the relationship between evolution and the genes
that regulate development became a field in itself: evolutionary development
or "evo-devo", for short.
As with the modern synthesis, itself, the unification of developmental biology and evolutionary biology occurred as scientists, learning new things in their own fields, recognized the relevance of their work to new areas. Consider the work of Jacob and Monod that we have already discussed, on the lac operon. Their work showed how genes could be turned on and off in response to environmental cues. Molecular biologists described ways that different genes were regulated in different cells: through the action of repressors and inducers (like transcription factors) that turn transcription on and off (often in response to external stimuli through signal transduction), through the differential splicing of m-RNA transcripts, through the differential splicing of initial protein products, and through the subsequent modification of those proteins. These are the processes that make cells in an organism different from one another during organismal development - they make bone tissue different from skin tissue, for example, and they determine the rate at which these tissues will grow. They regulate the growth of tissues and organs and the subsequent shape of an organism - its morphology. Evolutionary biologists and molecular geneticists realized that these regulatory genes, that influence the co-ordinated development of tissue specialization in organisms, might be the drivers of major evolutionary innovations. The second realization was that, because most animals share a similar embryological development, that these genes that regulate how development occurs should be largely homologous - even if the pathways that they regulate lead to the formation of very different (and non-homologous) structures.
 B.
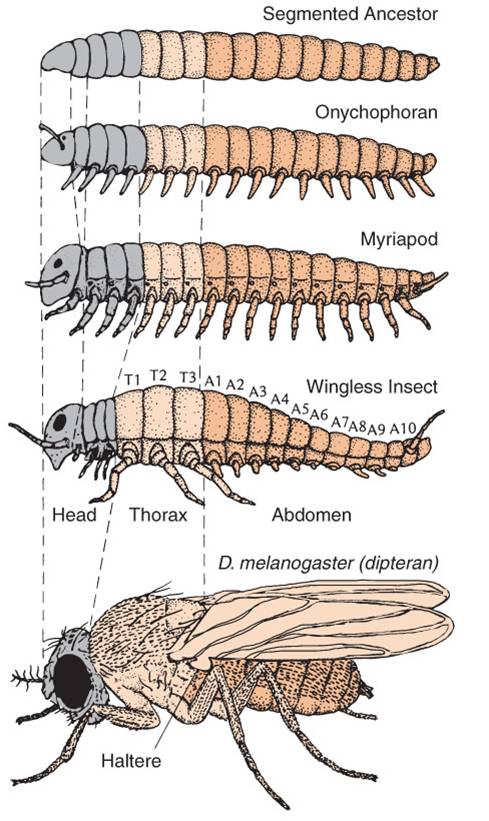
Homeotic Genes
B.
Homeotic GenesAs we will see when we get to animal diversity, almost all animals are 'segmented'. Like the segments on a worm, insect, or vertebrate spinal column, segmentation of body parts allows for specialization of structure and function. Much like gene duplication is a source of genetic novelty (keeping the original function while also allowing for the evolution of something new in the duplicated gene), dividing (or multiplying) the body into segments allows some segments to evolve new function without compromising the ability of the organism to do the 'old stuff', too. In one of the most remarkable and surprizing genetic discoveries of the last 30 years, molecular developmental geneticists were able to decipher how a group of genes act synergistically to coordinate the process of segmentation and subsequent specialization in the developing embryo. Although these genes and their activities were originally described in Drosophila fruit flies, the prediction of homology among animals for the genes regulating these fundamental processes was supported - these genes that regulate segmentation have been found in all segmented animals, from worms to vertebrates. In fact, ancestral homologs to these genes have also been found, in fewer number, in non-segmented Cnidarians (corals, jellyfish, and hydra).
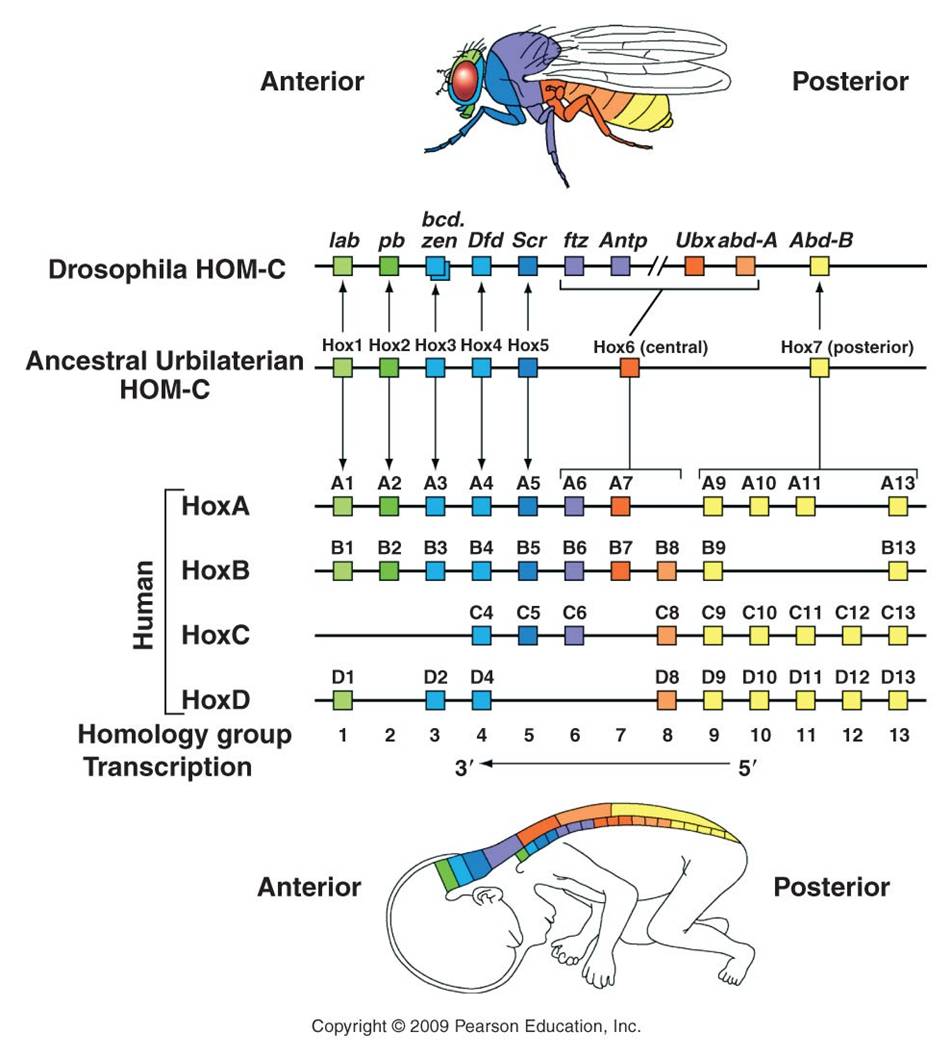
The relationship between gene duplication and segment specialization is even more direct. These homeotic genes that regulate development are very similar to one another, and have undoubtedly arisen by gene duplication. One family of genes all have a very similar sequence that codes for a DNA binding domain (as we would expect for a transcription factor that must bind DNA). This sequence is called a "homeobox", and the genes in this family are called the Hox genes. So, we see the process of duplication occurring at a genetic level, and causing body segment duplication at the organism level, with the same evolutionary consequence of providing the potential for new evolutionary innovation.
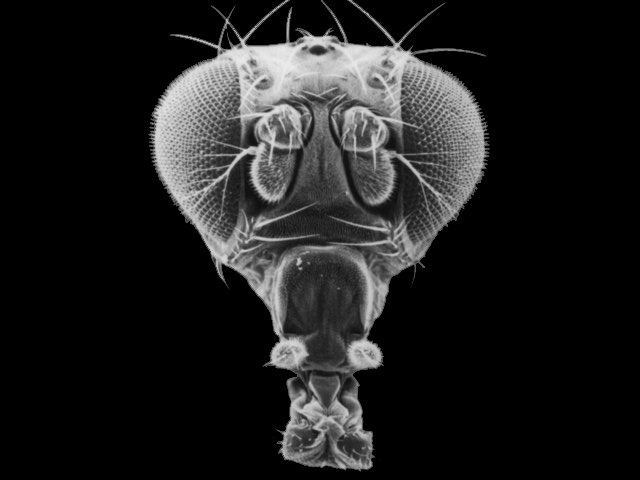
Changes in these genes can have profound effects on the final morphology of a segment. And because these genes encode transcription factors that are binding to MANY genes in the same cell, there are correlated and co-ordinated responses among the genes in a cell and the cells in that segment. Consider the "antennapedia" gene in Drosophila. It is "on" in the thoracic segments of a developing fly pupa during metamorphosis. The gene product - a transcription factor - stimulates the production of myriad genes that leads to the development of muscle, nerve, and exoskeleton tissue in the shape of a leg. This gene is normally off in head and abdominal segments, so legs don't develop there. However, mutants occur that express this gene in the head segment that usually gives rise to the antennae. In these individuals, the activation of the gene causes the development of legs on the head - where antennae should be.
 |
 |
 |
| Normal fruit fly, with normal antennae on front of head | Mutant, with legs instead of antennae | The effect of the 'Ultrabithorax' gene, on right. |
Another important hox gene in Drosophila, homologous to antennapedia and next to it (both suggesting that it is a gene duplicate) is the "ultrabithorax" (Ubx) gene. This gene codes for a m-RNA transcript that is spliced 5 different ways, and so is translated into five different transcription factor proteins. Normal expression of Ubx in the third thoracic segment of the fly interrupts development of a wing, and stimulates development of a larger set of legs on the third thoracic segment. Mutation in Ubx in this segment makes this thoracic segment develop like the second thoracic segment, growing wings and a smaller pair of legs.
Vertebrates have homologs of these same homeotic genes, with different numbers of copies of each that have arisen by duplication and deletion in the vertebrate lineage (see the colorful figure, above). Now, vertebrates don't have antennae or membraneous wings. However, these regulatory pathways still exist that turn on different sets of genes in these animals. These homeotic genes are like "genetic switches". Think about an electronic switch - it can flip on an incandescent light, a flourescent light, a blender, or a TV. The key is that the switch is linked to a given circuit, or process. One of the most dramatic examples of this 'switch' analogy involves the homeotic gene for eye development that occurs in all bilaterally symmetrical animals, including fruit flies and humans. If the gene is cut out of a developing fly embryo, then its head segments lack the gene and the fly does not produce eyes. However, if the human version of the switch is inserted in these mutants, the resulting flies develop eyes. Now, they don't develop VERTEBRATE eyes (thank goodness!), they develope the compound eyes of insects. So, the only thing the human gene did was to restore the switch, re-establishing the 'circuit' of eye development. Although the downstream effects are different (like turning on an incandescent light or florescent light), the switch is the same. A biological process can be linked to a homeotic switch by associating a gene with a binding site for that transcription factor - now that gene will be turned on when that transcription factor is present in the cell. Remember the picture of the promoter/operator region in the metallothionein IIA in humans? Remember how many different binding sites there were for different trancription factors? That gene could regulated by a variety of factors, and incorporated into a variety of metabolic/genetic processes. So, the organism's metabolism would change WITHOUT a change in the actual sequence of the metallothionein gene. Rather, there has been a change in how it is regulated.
 C.
Environmental Effects and Phenotypic Plasticity
C.
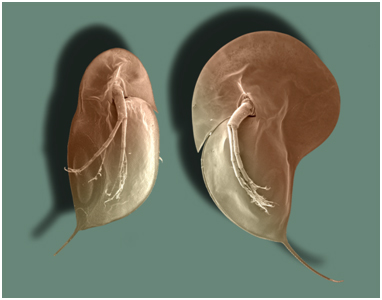
Environmental Effects and Phenotypic PlasticityThrough signal transduction, the changes occuring in one cell can affect other cells. Likewise, substances in the environment can affect gene expression - even the expression of developmental genes. For example, small freshwater crustaceans called Daphnia will grow larger spines and a larger head capsule in the presence of predatory fish. In the picture to the right, the Daphnia on the right grew in the presence of fish, and grew a bigger head capsule and larger posterior spine than the Daphnia at left, which grew in the absence of fish. So, a single genotype can produce a range of phenotypes - not just one. This is called "phenotypic plasticity", and the range of phenotypes that a genotype can produce are called its "reaction norm". As such, a phenotype is not a "hard-wired" response... rather, it represents a range of potential responses, with the observed response determined, in part, by environmental influences. This allows a population to 'experiment' with a wider range of phenotypes in a given environment - more variation upon which selection can act. Initially, a variant that is not genetically hard-wired can become encoded - as the genotype evolves to produce that phenotype more efficiently. Indeed, many genotypes may be producing the same adaptive phenotype by different genetic means - which maintains genetic variation in the population and maintains the ability to respond to still more changes in the environment. So again, evolution may not be driven through the production of new genes, but rather by the timing and coordination of gene interaction, under the influence of particular environmental cues.
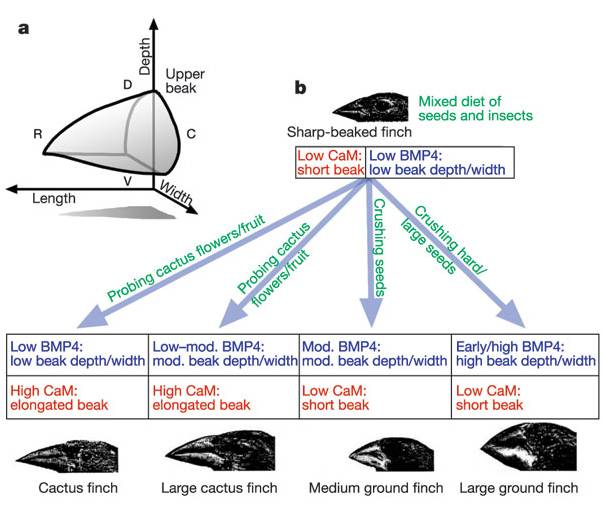
The action of regulatory genes can certainly create phenotypic variation that causes differential reproductive success (selection). Consider beak shape in Darwin's finches - one of the iconic examples of adaptive variation. How did these birds evolve different beak shapes? We can appreciate that different beaks would be adaptive in different environments with different food resources, but how are differences in beak shape encoded? It is not in the type of genes that are present, but rather in the timing and amount of their activity. In 2006, Abzhanov and coauthors described the effect of two interacting genes on beak shape. The first is BMP-4 - a "bone morphogen protein" - a transcription factor that stimulates collagen production and thus the growth of bone and cartilage. All Darwin's finches have this gene - the difference is that in large ground finches, the gene is turned on earlier in development and stays on longer than in the other species. Experiments with chicken embryos revealed that more of this protein results in larger beak development. A second gene produces calmodulin - which regulates calcium signalling in cells. High activity of this gene results in a pointy beak. Different combinations of these regulatory proteins results in different shaped beaks.
 |
 |
| The combined effects of two growth regulators, BMP-4 and calmodulin, influence the shape of the beak in species of Darwin's finches. | These differences in beak size and shape correlate with finch diet and ecological niche, and are believed to be important in the radiation and speciation occuring within this group. |
 E.
Allometry
E.
AllometryWhen we look at the diversity within a group of similar animals, what we usually see are changes in the proportion of different body parts - not really changes in the the types of parts, themselves. So, in mammals, we see the same skeletal system; the differences between mice and bats is not really in the types of bones they have, but rather in the relative sizes of the bones. So, the finger bones in a bat wing are long relative to the limb bones; whereas in mice, the finger bones are short relative to the limb bones. These differences occur because of the differences in the rates of growth of these bones during embryogenesis, and they are caused by changes in the activity of developmental genes. These changes in the relative growth of different body parts is called allometry. When we look at any large group or organisms - such as the insects or the vertebrates, what we see are differences on a single body theme - differences in shape due to relative differences in the sizes of the body parts. We now know that these differences in size, so important to creating the diversity of species that we see within a group - are largely do to differences in development. Again, the most remarkable discovery in evolutionary biology over the last 30 years is the discovery that differences between large groups of organisms - like the differences between insects and vertebrates - occurs despite an extraordinary similarity in the developmental processes and developmental genes that regulate their development.
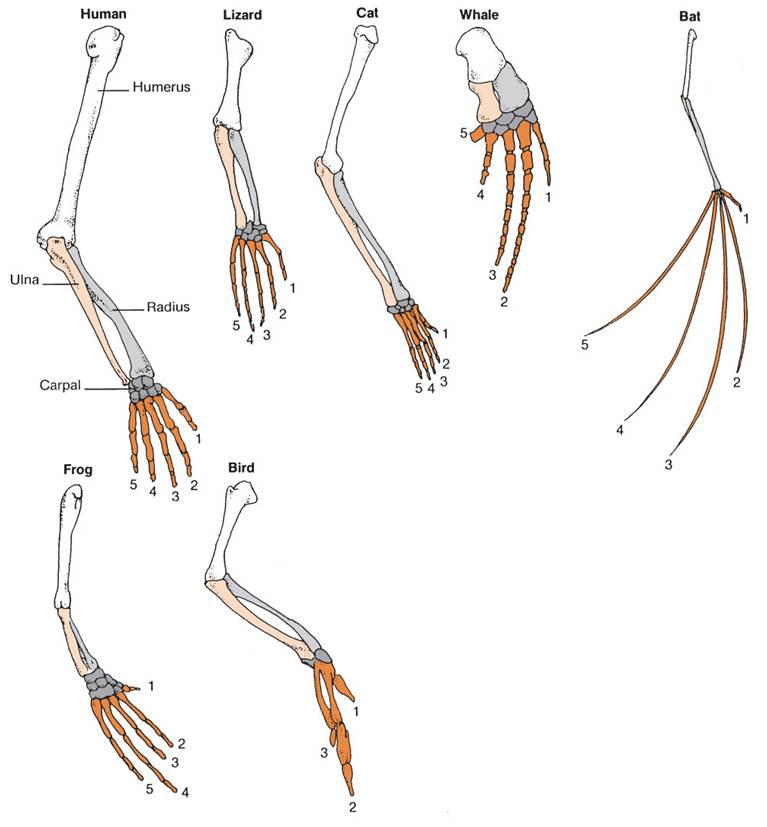
Likewise, homologous structures in a large group of organisms, like limbs in terrestrial vertebrates, are NOT the result of changes in structural genes. These structures are built out of the same material. Rather, the differences in the relative proportions of these bones is due to differences in developmental genes that alter the relative growth rates of the bones in the limbs and digits. Of course, the changes in the frequencies of these genes can be affected by our five agents of evolutionary change. In particular, as we saw with finch beaks, these changes in the proportions of body parts can be as adaptive as the genes for structural proteins like flower color. So, although the modern synthesis described a model of microevolutionary changes in structural genes within a population, the elucidation of developmental genes that influence to large scale structure of body plans allowed evolutionary biology to explain changes in body plan with the same model - but as a change in the gene frequencies of different genes - the genes that influence the process and pattern of development, itself. So, 'macroevolutionary' differences are the result of 'microevolutionary' processes occurring in developmental genes; 'macroevolution' and 'microevolution' are not two different things.
For another example, consider one genus of Scarab beetles, the genus Onthophagus. The different beetle species in this genus have a wide variety of horn shapes and sizes; the males use the horns to battle for mates (intrasexual selection), and they may also be under some control by intersexual selection (preference of females for males with a particular horn shape). If the ratio of horn size to body size is calculated for many individuals within these species, the species cluster in a manner consistent with their hypothesized phylogenetic relationships. So, it would seem that allometric growth in horn size has been an important factor correlating with divergence in this genus.
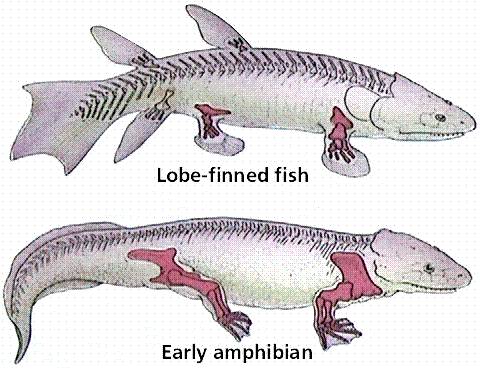
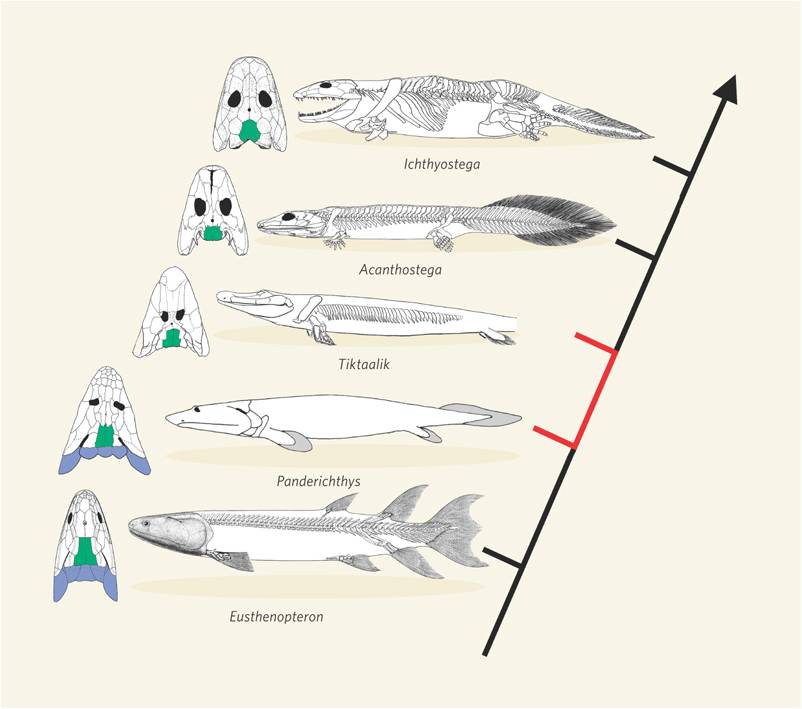
And, for yet another example, consider the evolutionary relationships between lobe-finned fishes and amphibians. There is now a very nice sequence of intermediate fossils of transitional proto-amphibians in the fossil record. When the structure of the bones in lobe-finned fishes, transitional fossils, and amphibians is examined, the changes are not in the presence of new bones, but rather in the spatial relationships, relative sizes, and developmental patterning of limb architecture. These changes were probably driven by changes in regulatory genes and transcription foactors.
 |
 |
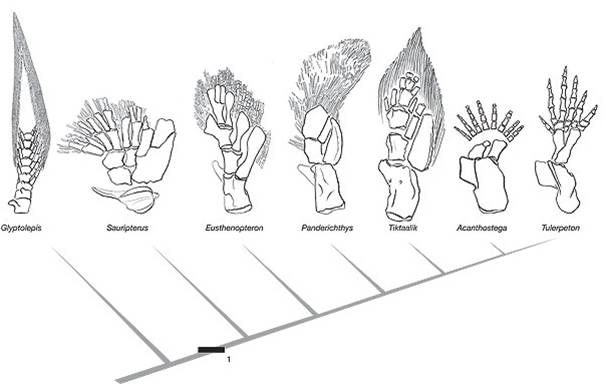 |
| The limb bones in lobe finned fishes and terrestrial vertebrates are homologous. | The transition from fish to terrestrial amphibians is well documented in the fossil record | The change in forelimb structure is really one of a change in the relative position of skeletal units and skeletal architecture. |
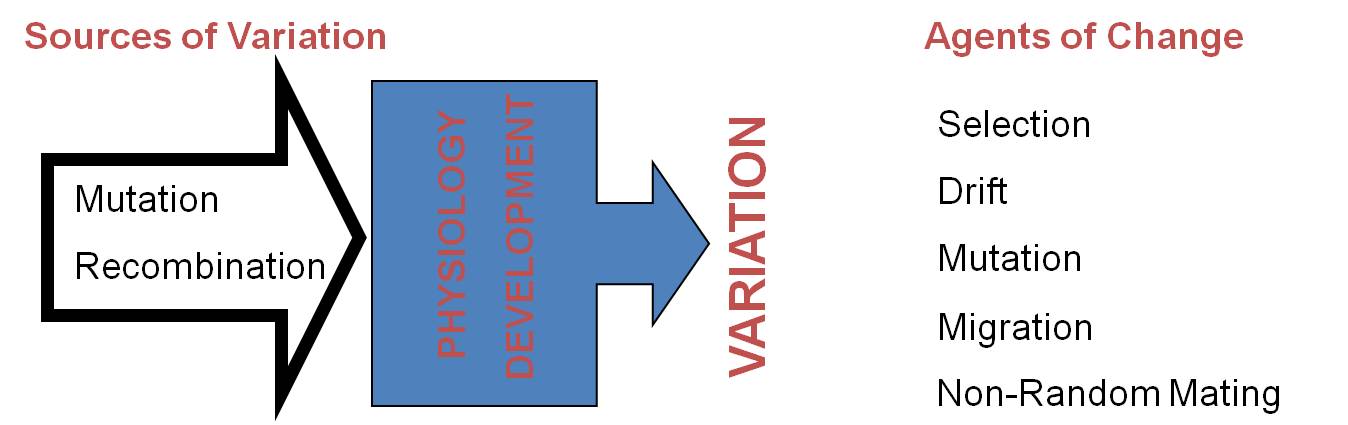 Large
scale differences in morphology that define the major groups of organisms -
which are sometimes refered to as "macroevolutionary" patterns - are
probably not caused by changes in structural genes. However, this does not mean
that these changes are orchestrated by different processes. Rather, these macroevolutionary
changes that occur in large-scale morphological characteristics are probably
caused by changes in the action of regulatory genes like transcription factors,
which affect developmental processes. The changes in the action of these genes
can be modeled by population genetics, too; and we would expect the effects
of the genes to be acted on by natural selection, genetic drift, positive assortative
mating, mutation, and migration. So, in this sense, microevolutionary changes
that occur within a population at a given locus can cause the macroevolutionary
patterns that we see in the evolution of new, different types of organisms.
It just that those microevolutionary changes in gene frequencies are occurring
at the loci of regulatory genes that co-ordinate morphological development in
that population.
Large
scale differences in morphology that define the major groups of organisms -
which are sometimes refered to as "macroevolutionary" patterns - are
probably not caused by changes in structural genes. However, this does not mean
that these changes are orchestrated by different processes. Rather, these macroevolutionary
changes that occur in large-scale morphological characteristics are probably
caused by changes in the action of regulatory genes like transcription factors,
which affect developmental processes. The changes in the action of these genes
can be modeled by population genetics, too; and we would expect the effects
of the genes to be acted on by natural selection, genetic drift, positive assortative
mating, mutation, and migration. So, in this sense, microevolutionary changes
that occur within a population at a given locus can cause the macroevolutionary
patterns that we see in the evolution of new, different types of organisms.
It just that those microevolutionary changes in gene frequencies are occurring
at the loci of regulatory genes that co-ordinate morphological development in
that population.
Study Questions:
1. What is epigenesis?
2. Explain the relationship between development and evolution.
3. How are gene duplication and segmentation similar in terms of introducing evolutionary novelty?
4. What are homeotic genes, and what type of protein do they code for?
5. Explain homologous structures in terms of the regulation of developmental genes.