 We
have summarized some of the major epochs in the history of the Earth before,
but we will review them here and add in important evolutionary events in the
history of life.
We
have summarized some of the major epochs in the history of the Earth before,
but we will review them here and add in important evolutionary events in the
history of life. The use of fossils and genetic phylogenies have allowed scientists to reconstruct the history of life and the major epochs and innovations of evolution that have occurred over the last 4 billion years of earth's history. Perhaps the most surprizing thing that we have learned is that we humans are very late arrivals on the evolutionary stage, and that the play had been going on just fine for quite a long while without us. Indeed, the evolution of anything you might recognize as an 'animal' is a fairly recent event in the evolutionary history of the planet; animals have only been present for the last 1/7th of earth history. Although we might think of macroscopic animals and plants as the 'most important' forms of life somehow, it is really the little things - bacteria and archaeans - that keep the biosphere running and allow animals and plants to survive. Without bacteria, you would not digest your food efficiently, plants would have a more difficult time absorbing nutrients, and the nutrients in dead organisms would not be recycled and made available to those of us that are still alive. The basic life processes that drive the biosphere - things like nitrogen fixation, photosynthesis, respiration and energy conversion, and decomposition - are still driven by bacteria. In the first part of this lecture, we'll see that is, indeed, how it has been for the majority of earth history.
We
have summarized some of the major epochs in the history of the Earth before,
but we will review them here and add in important evolutionary events in the
history of life.
4.5 billion years ago - Earth formed (Evidence: meteorites and moon rocks date to this age, dating the condensation of planetissimals in the solar system)
4.0 bya - oldest rocks (The oldest rocks are not as old as the Earth because the Earth's surface was molten - as a result of constant meteorite bombardment and the energy released during the stratification of Earth's core, mantle, and crust. In addition, the Earth is tectonically active, recycling its crust. So, older rocks that may have solidified may have been subducted and melted)
3.5-3.8 bya - origin of life (The oldest fossils, both microfossils and stromatolites, date to 3.5-3.8 bya). These organisms are similar to prokaryotes we see today. However, it is not possible to distinguish whether they are Bacteria or Archaeans. However, from this period forward there are anaerobic and non-oxygenic photosynthetic bacteria.
2.5-2.0 bya - oxygen accumulating ("red bed" sediments form, demonstrating that oxygen gas in the water was oxidizing iron); this is evidence of the evolution of oxygenic photosynthesis. With the accumulation of oxygen, many obligate anaearobes undoubtedly go extinct. However, other bacteria evolve the capacity to USE oxygen as an oxidizing agent, making the harvest of energy from glucose metabolism more efficient... in other words, aerobic respiration evolves.
2.0 bya - eukaryotes evolve through endosymbiotic absorption/tolerance of aerobic bacteria (protomitochondria). With the evolution of eukaryotes comes the evolution of sexual reproduction. These endosymbionts may have been a cellular parasite, like Rickettsia. Subsequent lines of eukaryotes absorbed photosynthetic cyanobacteria, which evolved into symbiotic chloroplasts. The evidence for these theories of eukaryote evolution are the genetic similarity between mitochondria and chloroplasts (that have their own DNA) and various types of bacteria and cyanobacteria. So until this point - representing nearly 1/2 of life's history on this planet - life was exclusively bacterial. The major types of ecological roles, from nitrogen fixation, to photosynthesis, to heterotrophy, to saprophagy (eating dead stuff) all evolved among bacteria - creating food webs and biogeochemical cycles that shunted energy through the biosphere and other earth systems.
0.7 bya - the first animals evolve - sponges and then soft-bodies invertebrate fossils, often radially symmetrical like jellyfish.
0.5 bya - radiation of animals with hard bodies (Cambrian explosion) - beginning of the Paleozoic Era ('old animals')
0.4 bya - evolution of land animals and plants
0.24 bya (240 mya) - Permian extinction; end of the Paleozoic Era
240-65 mya - Mesozoic Era (Age of Dinosaurs - 'middle animals')
65 mya - Cretaceous extinction (meteorite impact off Yucatan Peninsula) - beginning of the 'Cenozoic Era' ('recent animals')
6-5 mya - split of chimp and human lineages
4.5 mya - Australopithecines (1/1000 of earth's history left to present...99.9 % has occurred already).
0.2 mya (200,000) - oldest Homo sapiens fossil. (99.995 % of earth's history has passed).
The Linnaean system is a heirarchical system that places species within genera, and similar genera within higher nested categories. Historically, these determinations were based on similarities in morphology. There are lots of mnemonics for remembering the Linnaean classification system: "Dandy King Philip Came Over From Great Spain" is one way to remember the sequence of "Domain, Kingdom, Phylum, Class, Order, Family, Genus, and Species". However, it doesn't work for plants, in which 'Division' is used instead of phylum. The Linnaean system lended itself to evolutionary reinterpretations, because evolution also produces nested patterns of relationships. The process of divergence creates an evolutionary 'tree', and each twig (species) is nested with other small twigs as part of one branch, and several branches form one limb, and the limbs unite to form the trunk.
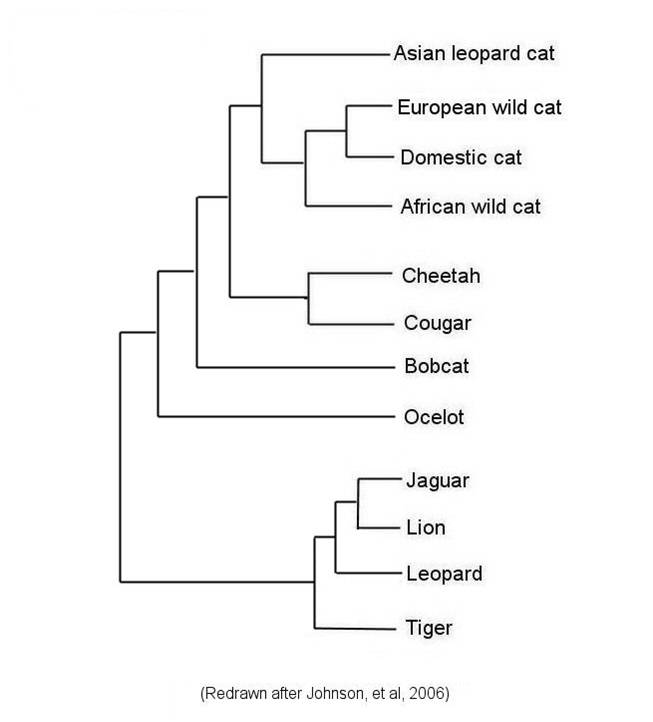
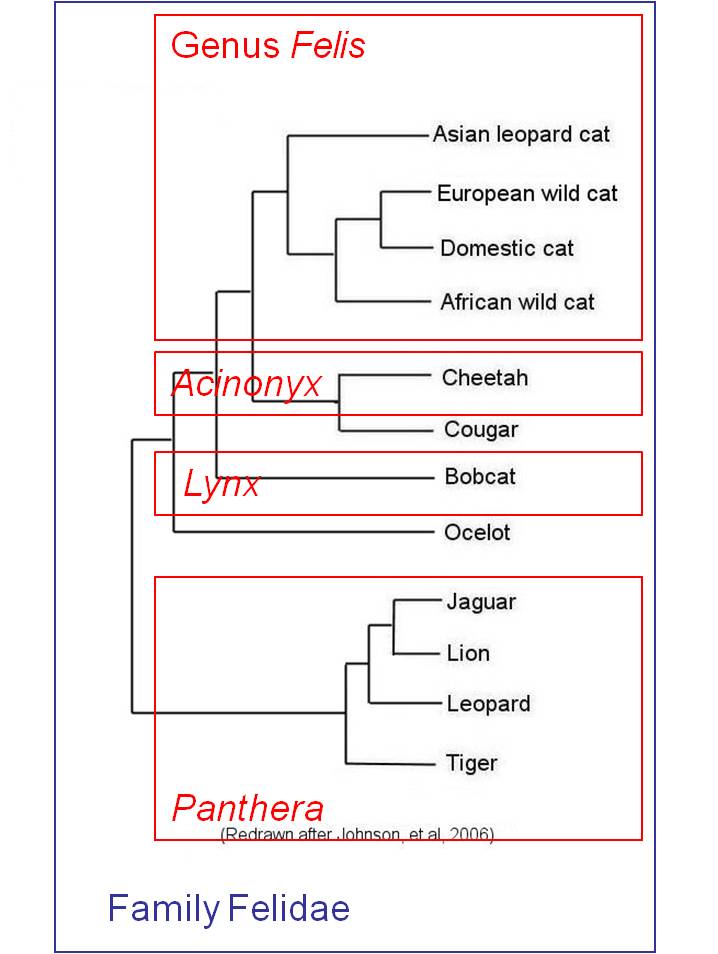
Although Linnaean classification and evolution both produce nested heirarchies, they are not necessarily equivalent. For instance, if we have multiple genera within a family, the Linnaean system does not necessarily describe the relationships among these genera; all it describes is their mutual presence in this one family. For example, in the figure below left, the Linnaean systems allocates cats into four genera, but does not describe the relationships among these genera. However, evolution - through phylogenetic systematics - tries to determine which taxa (species, genera, families, etc.) are more closely related and share a more recent common ancestor. Indeed, phylogenetic systematics would argue that the degree of biological relatedness among taxa should be reflected in their taxonomic affinity. This becomes a goal of phylogenetic systematics - to create a taxonomy that represents the evolutionary relationships among species, genera, families, etc. The goal is to create a monophyletic taxonomy that include all organisms decended from a common ancestor.
 |
 |
 |
 The
Linnaean system was based on morphology, alone, and we know that morphology
can sometimes misrepresent patterns of genetic relatedness (remember sibling
species and polymorphic species?). So, as we learn more about the degree of
genetic relatedness among species, we occasionally find that some species that
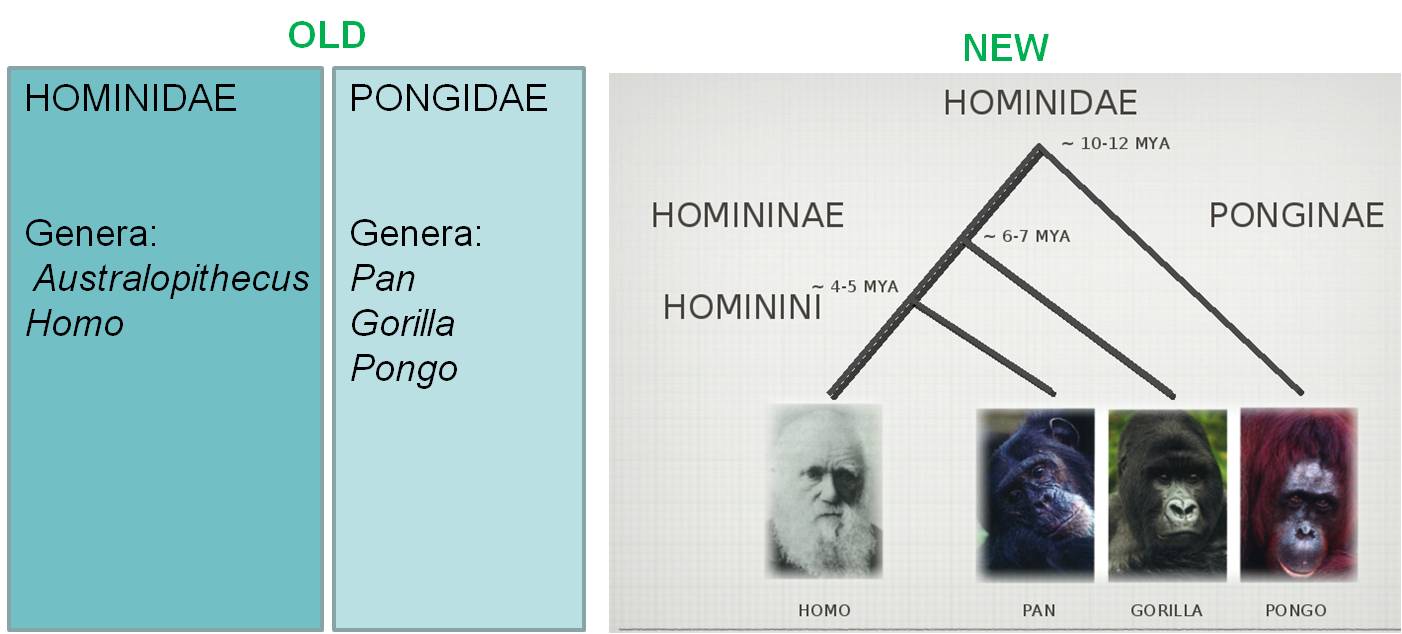
are closely related have been placed in different taxa. For example, the historical
classification of humans, chimps, gorillas, and orangutans placed humans in
the family Hominidae, and the other three species in the family Pongidae. This
does not represent their degree of biological relatedness, however, because
human and chimps share a more recent common ancestor with one another. So, to
include all descendants of a common ancestor (and that ancestor), chimps and
humans should be placed in the same taxonomic group (either genus or family).
This could include gorillas (as all three share a common ancestor), or could
also include gorillas and orangutans (as all four share a common ancestor),
but can't split humans and chimps into separate groups.
The
Linnaean system was based on morphology, alone, and we know that morphology
can sometimes misrepresent patterns of genetic relatedness (remember sibling
species and polymorphic species?). So, as we learn more about the degree of
genetic relatedness among species, we occasionally find that some species that
are closely related have been placed in different taxa. For example, the historical
classification of humans, chimps, gorillas, and orangutans placed humans in
the family Hominidae, and the other three species in the family Pongidae. This
does not represent their degree of biological relatedness, however, because
human and chimps share a more recent common ancestor with one another. So, to
include all descendants of a common ancestor (and that ancestor), chimps and
humans should be placed in the same taxonomic group (either genus or family).
This could include gorillas (as all three share a common ancestor), or could
also include gorillas and orangutans (as all four share a common ancestor),
but can't split humans and chimps into separate groups.
Another classic example is the treatment of mammals, birds, and reptiles. In Linnaean taxonomy, these three groups of animals are placed in different Classes: Mammalia, Reptilia, and Aves (birds). But phylogenetic systematics groups them differently (see the tree of life site). Some reptilian groups, like extinct dinosaurs and extant crocodilians, are more closely related to birds than they are to other reptiles like lizards, snakes, and turtles. And some "reptiles", like the sail-finned Pelycosaurs, are more closely related to mammals than to other reptiles. So, phylogenetic systematics separates the "synapsids" (mammals and mammal-like reptiles), from "reptilia", and divides reptilia into the "anapsids" (turtles) and "diapsids". The diapsids are further divided into the "archosauromorphs" (crocodilians, birds, and extinct dinosaur relatives) and the "lepidosauromorphs" (modern lizards ands snakes, and their extinct ancestors). Because the number of clades does not conform to the number of taxonomic levels, the notion of "class" and "order" becomes problematic. However, knowing that "archosauromorphs" contains croc and birds but not turtles, lizards, or snakes correctly represents the evolutionary relationships among these groups, and conveys more correct evolutionary information than the term 'reptile'.
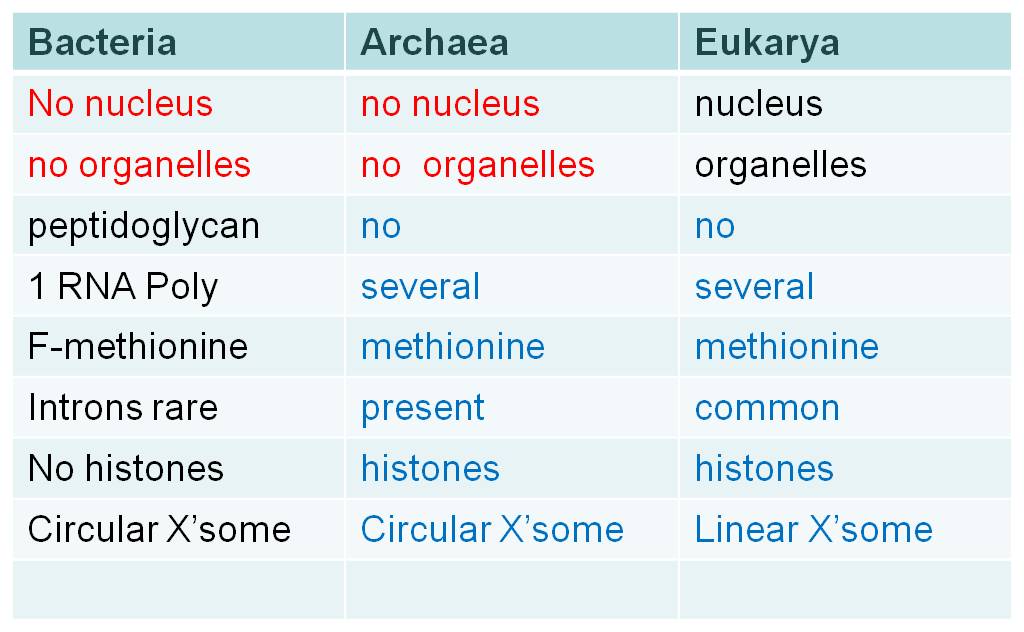
Finally, a third example highlights the issue at the largest scale of taxonomic resolution, and brings us back to bacteria. Prior to the 1980's, most biologists classified all prokaryotes in the Kingdom Monera. While they recognized that there was great diversity in the group, biologists considered the absence of a nucleus a fundamental, distinguishing morphological feature of the group that separated it from all eukaryotes. However, genetic analyses in the 1980's by Carl Woese suggested that there were two major clades of prokaryotes - the Bacteria and the Archaea. And actually, most genetic analyses show that the Archaea are more closely related to the eukaryotes than to the bacteria. So, using the criteria of phylogenetic systematics, it is improper to classify all prokaryotes together. Rather, Woese suggested that a new level be created - the domain, and that life be split into three domains: the Bacteria, the Archaea, and the Eukarya.
III. The Prokaryote Domains: Bacteria and Archaea
A. Overview
 These
two domains are ancient; they may even have diverged before 3.5 bya. The problem
with using genetic clocks and genetic similarity to attempt to date this point
of genetic divergence on these groups is that prokaryotes exchange DNA "laterally".
They don't just pass it down a lineage from parental cell to daughter cells;
they can also pass genes from one living bacterium to another through conjugation
or transformation. As such, genetic similarity can occur as a result of lateral
gene transfer between unrelated organisms, and thus the degree of genetic similarity
in particular genes does not necessarily indicate the degree of common ancestry.
For these reasons, it is very difficult to use this particular technique to
date the time of divergence of these two ancient groups.
These
two domains are ancient; they may even have diverged before 3.5 bya. The problem
with using genetic clocks and genetic similarity to attempt to date this point
of genetic divergence on these groups is that prokaryotes exchange DNA "laterally".
They don't just pass it down a lineage from parental cell to daughter cells;
they can also pass genes from one living bacterium to another through conjugation
or transformation. As such, genetic similarity can occur as a result of lateral
gene transfer between unrelated organisms, and thus the degree of genetic similarity
in particular genes does not necessarily indicate the degree of common ancestry.
For these reasons, it is very difficult to use this particular technique to
date the time of divergence of these two ancient groups.
Although they share some fundamental similarities like the lack of a nucleus, the lack of membrane-bound organelles, and circular chromosomes, these two prokaryotic domains differ in some important respects. When they differ, the archaeans are often similar to the Eukarya. For example, bacteria have peptidoglycan in their cell walls, while archaeans (and eukaryotes) do not. Bacteria have only one type of RNA Polymerase; archaeans (and eukaryotes) have several. Bacteria use formyl-methionine as the initiator amino acid in translation, archaeans (and eukaryotes) use methionine. Histones are not associated with bacterial chromosomes, but they are associated with archaean (and eukaryote) chromosomes. Introns are rare in bacterial genes, but are progressively more common in archaeans and eukaryotes. So, there are several fundamental differences between these two groups, and often the archaeans are more similar to eukaryotes than to bacteria.
1. Archaea
 The
archaea are very diverse, and include groups that have adapted to some of the
most inhospitable environments on Earth. Indeed, many of these environments
may be similar to those that were common in early earth history. So, rather
than thinking that these organisms have evolved to exploit these isolated odd
habitats, perhaps they adapted to these conditions when the conditions were
common, and have just 'hung on' in these habitats that have become rare over
time as the earth has changed. The extreme thermophiles
live at very high temperatures at sulphur springs and geothermal vents where
temperatures can exceed 1000C. How they protect their proteins and
DNA under these conditions is an area of active research. Another group are
tolerant of very extreme salt concentrations - these are the extreme halophiles.
Yet another group of extreme organisms are the methanogens. These organisms
use H2 to reduce CO2 and produce energy; producing CH4
(methane or 'swamp gas') as a waste product. Most are obligate anaerobes, meaning
that they can only survive in environments where the concentration of oxygen
gas is very low. However, some are aerotolerant, and can survive in the presence
of oxygen though they do not use it for respiration. These conditions occur
at the bottom of deep water bodies, or where the action of respiring bacteria
deplete available oxygen - like in swamp and marsh sediments and in the digestive
tract of animals (including humans). Some methanogens are extreme thermophiles,
too.
The
archaea are very diverse, and include groups that have adapted to some of the
most inhospitable environments on Earth. Indeed, many of these environments
may be similar to those that were common in early earth history. So, rather
than thinking that these organisms have evolved to exploit these isolated odd
habitats, perhaps they adapted to these conditions when the conditions were
common, and have just 'hung on' in these habitats that have become rare over
time as the earth has changed. The extreme thermophiles
live at very high temperatures at sulphur springs and geothermal vents where
temperatures can exceed 1000C. How they protect their proteins and
DNA under these conditions is an area of active research. Another group are
tolerant of very extreme salt concentrations - these are the extreme halophiles.
Yet another group of extreme organisms are the methanogens. These organisms
use H2 to reduce CO2 and produce energy; producing CH4
(methane or 'swamp gas') as a waste product. Most are obligate anaerobes, meaning
that they can only survive in environments where the concentration of oxygen
gas is very low. However, some are aerotolerant, and can survive in the presence
of oxygen though they do not use it for respiration. These conditions occur
at the bottom of deep water bodies, or where the action of respiring bacteria
deplete available oxygen - like in swamp and marsh sediments and in the digestive
tract of animals (including humans). Some methanogens are extreme thermophiles,
too.
Although the Archaea are noted for representatives that tolerate these extreme environments, there are other archaeans that live under more benign environmental conditions across our planet, where they function as photosynthetic producers, consumers, and decomposers. Curiously, although archaeans do inhabit the human gut, no archaeans have been identified that cause human disease. The other prokaryotes - the bacteria - contain many species that cause human disease.
2. Bacteria
 The
bacteria are as diverse than the archaeans, expressing an extraordinary variety
of metabolic processes. A couple notable groups are the cyanobacteria, thought
to share common ancestry with eukaryotic chloroplasts, and the gram-positive
bacteria that have many types of pathogenic bacteria, such as those causing
staph and strep infections, botulism, tuberculosis, anthrax, and many other
diseases.
The
bacteria are as diverse than the archaeans, expressing an extraordinary variety
of metabolic processes. A couple notable groups are the cyanobacteria, thought
to share common ancestry with eukaryotic chloroplasts, and the gram-positive
bacteria that have many types of pathogenic bacteria, such as those causing
staph and strep infections, botulism, tuberculosis, anthrax, and many other
diseases.
Both the bacteria and archaea are very old groups, and have been radiating and evolving for over 3.0 billion years. Recent genetic studies have revealed an astounding degree of genetic variation within these groups - an understandable consequence of their long evolutionary histories. This genetic variation is reflected by the 'branch lengths' in the tree diagram to the right. The genetic variation among the plants, animals, and fungi, represented by their relative short branch lengths, are dwarfed by the genetic variation among the archean and bacterial clades. But of course, the animals have only been radiating for the last 0.7 billion years. So, the archean and bacterial lineages have been radiating for 5 times as long and so it is not surprizing that they are much more diverse. Thus, as expected for groups that are so old, they have radiated dramatically - even within subgroups. For this reason, even groups that share common ancestry can include organisms that are very different. For our purposes here, an overview of the diversity is presented, rather than a overview of their classification or phylogeny.
If we examine the morphological diversity in prokaryotes, we find round (coccus), rod-shaped (bacillus), and spiral (spirochaete). That is not a lot of morphological diversity! So how have the prokaryotes been changing over the last 3.5 by? Well, they have been evolving metabolically. The theme of bacteria diversity is metabolic diversity. There are very few metabolic processes that eukaryotes can perform that prokaryotes cannot. However, the reverse is not true; the prokayotes have a wide range of metabolic diversity - a range that, in many respects, exceeds the metabolic diversity of eukaryotes.
1. Requirements of Oxygen for Cellular Respiration
Consider types of respiration. There
are no anaerobic eukaryotes. Most eukaryotes require oxygen (they have mitochondria,
after all), although some can function without it for a short time (facultative
anaerobes like yeast). Prokaryotes, on the other hand, have a wide range of
'respiratory' strategies:
- some bacteria, like methanogens, cannot live in the presence of oxygen - obligate
anaerobes
- some bacteria can tolerate oxygen
but don't use it - aerotolerant
- some bacteria can live with or without oxygen - facultative aerobes
- some bacteria live only in the presence of oxygen - obilgate aerobes
2. Metabolic Pathways for Harvesting Energy and Carbon from the Environment
1. Chemolithotrophs:
Use H2S or NH3 as an electron donor to power ATP production without sunlight;
and then use ATP to synthesize glucose from CO2. So, they harvest their
own energy from chemicals and make their own large organic molecules.
This is only done by bacteria.
2. Photoautotrophs:
Use either H2S (non-oxygenic) or H2O (oxygenic) photosynthesis powered by sunlight
to make ATP. Use the ATP to make glucose. So, they harvest energy
from light and make their own carbon resource. Cyanobacteria are oxygenic, and
their ancestors are probably responsible for the oxygen revolution about 2.5-2.0
bya. In addition to bacteria, some protists (algae) and most plants use this
metabolic strategy. The genetic similarity between chloroplasts (in algae and
plants) and certain cyanobacteria suggest that cyanobacteria may be ancestral
to chloroplasts,having been engulfed by phagocyctosis billions of years ago.
3. Photoheterotrophs: These strange organisms use sunlight as a source of energy for the formation of ATP, but they do not synthesize their own large carbon molecules (glucose). Rather, they have to absorb these molecules from the environment (heterotrophic). This is only done by some types of bacteria.
4. Chemoheterotrophs - must get BOTH energy and carbon from the metabolism of complex organic molecules. This is the most common strategy for living systems, exploited by some bacteria, some protists, all fungi, and all animals.
3. Ecological Roles
- primary producers: bacteria still make a significant contribution to the absorption of CO2 and the production of oxygen on our planet - largely in the oceans. And these oceanic bacteria are the bottom of marine food chains.
- nitrogen-fixing bacteria: can convert N2 into NH3 (ammonia) and then NO3 (nitrates) which can be taken up by plants. Only bacteria can do this; and so all new, natural inputs of nitrogen to the biosphere are by bacteria. (Lots of biologically accessible nitrogen is added to the environment by humans - through fertilizer and the production of NOx compounds by internal combustion engines).
- decomposers: as consumers of dead organic matter, bacteria (and fungi) break down complex organics and harvest energy. And the byproducts of that digestion are mineral nutrients (N, P, K) that are made available for reuptake in the biosphere by plants. So, bacteria are needed for nutrients to cycle through ecosystems.
- pathogenic bacteria: these bacteria get energy from living tissue and destroy cells in the process; they also produce either exotoxins that they secrete, or endotoxins that are bound to the cell wall. Examples of these pathogenic bacterial diseases are: staph infections, salmonella, typhoid, cholera, scarlet fever, tetanus, tuberculosis, botulism, gonorrhea, and syphilis.
- symbiotes: there are gut bacteria in nearly all animals that assist with digestion and may be required for the production and absorption of certain metabolites.There are also bacterial symbionts in plants, protists, and fungi.
As such, bacteria interact in every way possible with other organisms, and form the 'hub' of the biosphere. In some ways, the evolution of life has been an "embellishment" of the bacterial ecosystems that dominated the planet for the first 2.3 billion years of life.
The take home message about prokaryotes is their metabolic diversity. For 1.8 billion years, life on Earth was SOLELY bacterial. How was it evolving? How was it responding to the environment? Well, it seems that life was responding metabolically, adapting by exploiting different environments and energy sources in different ways.
Study Questions:
1. How do Linnaean classification and cladistic phylogenetic analyses differ?
2. How has the way we classify human changed as a consequence of genetic analyses?
3. What are the three domains of life?
4. Why are aarchaeans and bacteria so much more genetically diverse than plants, animals, and fungi?
5. What are "extremophiles? Give an example.
6. All eukaryotes are aerobic. List four types of metabolism that prokaryotes have with respect to oxygen.
7. Describe the four major types of metabolism that are shown by prokaryotes.
8. Describe five ecological roles that prokaryotes play in the environment.