 The
Diversity of Life The
Diversity of Life
The
Diversity of Life The
Diversity of Life 5. Major 'Invertebrate' Phyla
a. Phylum Porifera: Sponges
b. Phylum Cnidaria: Hydra, anemones, corals, jellyfish
The "Bilateria"
Protostomes:
Deuterostomes:
l. Phylum Chordata: Tunicates, Cephalochordates, and Vertebrates
 The
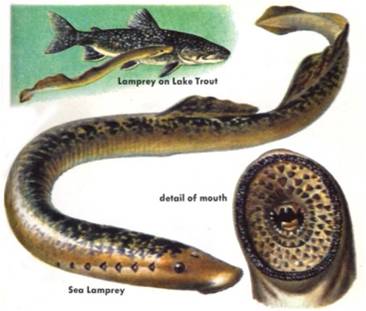
jawless fishes:
The
jawless fishes:
 The
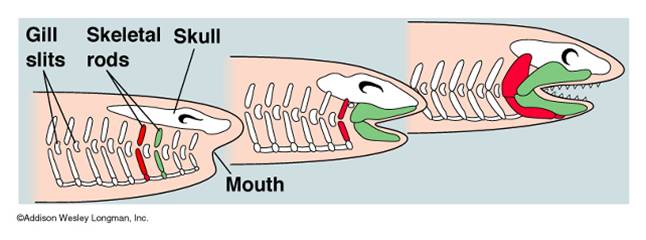
Gnathostomes:
The
Gnathostomes:
 The
Tetrapods: The
tetrapod clade includes all vertebrates descended from the first animals to
walk on land. The radiation of early tetrapods is very impressive; the adaptive
zone of the Carboniferous swamps was very large and well-suited to amphibian
animals reliant on water to lay their eggs. In addition, the huge amount of
biological productivity (lots of photosynthesis and less decompostion, remember?)
led to large populations of herbivorous insects that were eay prey for large
land vertebrates. The Carboniferous could well be called the "Age of Amphibians".
Many groups of early 'amphibians' radiate during this period, collectively called
the 'stem tetrapods'. However, the dry periods of the Permian and Mesozoic were
not optimal for these animals limited to areas near water. Today, three groups
of amphibians remain. The Caecilians are unusual, rare, legless amphibians.
Salamanders are the second group, and the giant Japanese salamander is the largest
living amphibian - reaching six feet in length! The most diverse group is the
Anura - mening "tailless" - the frogs and toads. Most amphibians have
gills or lungs at some point in their life cycle, but many also (or exclusively)
respire through their skin. For this to happen, the gases must diffuse into
a thin film of water, and then move across cell membranes by diffusion. So,
most amphibians need to keep their skin moist to respire; and it also means
that their skin, as an absorptive surface, is also prone to absorb toxins in
the environment. For these reasons, amphibians are particularly sensitive to
pollutants and environmental contaminants. Over the last 50 years, there has
been a dramatic decline in amphibian populations worldwide, probably due to
exposure to human-produced pollution, climate change, and chytrid fungal infections.
It is tough to say which of these variables is 'most' responsible for the decline.
In all likelihood, each factor increases sensitivity to the other two. In any
case, these three factors are having a decimating effect on amphibian populations.
The
Tetrapods: The
tetrapod clade includes all vertebrates descended from the first animals to
walk on land. The radiation of early tetrapods is very impressive; the adaptive
zone of the Carboniferous swamps was very large and well-suited to amphibian
animals reliant on water to lay their eggs. In addition, the huge amount of
biological productivity (lots of photosynthesis and less decompostion, remember?)
led to large populations of herbivorous insects that were eay prey for large
land vertebrates. The Carboniferous could well be called the "Age of Amphibians".
Many groups of early 'amphibians' radiate during this period, collectively called
the 'stem tetrapods'. However, the dry periods of the Permian and Mesozoic were
not optimal for these animals limited to areas near water. Today, three groups
of amphibians remain. The Caecilians are unusual, rare, legless amphibians.
Salamanders are the second group, and the giant Japanese salamander is the largest
living amphibian - reaching six feet in length! The most diverse group is the
Anura - mening "tailless" - the frogs and toads. Most amphibians have
gills or lungs at some point in their life cycle, but many also (or exclusively)
respire through their skin. For this to happen, the gases must diffuse into
a thin film of water, and then move across cell membranes by diffusion. So,
most amphibians need to keep their skin moist to respire; and it also means
that their skin, as an absorptive surface, is also prone to absorb toxins in
the environment. For these reasons, amphibians are particularly sensitive to
pollutants and environmental contaminants. Over the last 50 years, there has
been a dramatic decline in amphibian populations worldwide, probably due to
exposure to human-produced pollution, climate change, and chytrid fungal infections.
It is tough to say which of these variables is 'most' responsible for the decline.
In all likelihood, each factor increases sensitivity to the other two. In any
case, these three factors are having a decimating effect on amphibian populations.
The Amniota: Within the tetrapods, a new type of animal evolved in the Carboniferous; an animal that laid an egg surrounded by a series of membranes and a shell. These layers served to protect the developing embryo and yolk sac from dessication, allowing these species to colonize land further from water and exploit food resources and habitats that amphibians could not reach. This clade of animals is the amniotes, and their evolutionary innovation is the amniotic egg. The primitive condition within this group probably produced a leathery covering, much like turtles. A more rigid, desiccation resistant egg like birds have is probably a derived trait, as is the retention of the egg and live bearing of young, as seen in some snakes, and most mammals. The amniotes radiated during the Permian Period, when the drying of the unified land mass of Pangaea gave them a decided advantage over the ancestral, amphibious tetrapods. Three groups evolved at this time, distinguished by their skull morphology.
 The
Synapsids and the Mammals: The first group to dominate
were the synapsids. They dominated during the Permian, and were represented
by the great "sail-finned" Pelycosaurs and the Therapsid lineages
(the Gorgonopsians, the Dicynodonts, and the Cynodonts). The synapsid lineage
that is alive today are the Mammals. The transition from ancestral to derive
synapsids is well preserved in the fossil record, documenting the evolution
of the inner ear from the ancestral jaw bones, and the evolution of complex
and specialized dentition. The first true mammals are the Morganucodonts that
evolved in the Jurassic Period of the Mesozoic, about 200 mya. Ancestral synapsids
were large carnivores that dominated the landscape; a niche that would be filled
by the diapsid dinosaurs during the Mesozoic. The mammals were small, noctural
insectivores and scavengers. They radiated into these niches in the Jurassic,
perhaps because their endothermy allowed them to use the cool night more effectively
and under cover of darkness from larger predators. The surviving lineage of
this radiation are the three species of monotremes alive today: the platypus
and two species of echidna. These mammals lay eggs. Their offspring hatch, and
then nuzzle the belly of their mothers. Their mothers have sweat glands that
have evolved to produce a nutrient rich milk - they are mammary glands. However,
even the modern representatives lack nipples on these glands - so they "sweat"
milk onto their belly and the the hatchlings lick it up. Placental and marsupial
mammals evolved later, in the Cretaceous Period. Marsupials do not lay eggs;
they give live birth to a very small, embryonic newborn. The newborn climbs
from the genital opening to the pouch - a flap of skin covering the nipples
of the mammary glands. The embryo attaches to a nipple and feeds nearly continuously,
completing development in the pouch. This allows the mother to carry the embryo
with her for a longer period; there are no eggs that must be left periodically
for the mother to feed. The last group of mammals, the placentals, also retain
the embryo and give live birth. However, they retain the embryo much longer,
often until the offspring is capable of independent activity. The efficient
feeding of the embryo is facilitated by the placenta - which allows nutrients
in the bloodstream of the mother to pass directly into the bloodstream of the
offspring. This is far more efficient, energetically, than the mother turning
the nutrients into milk, that must be consumed and digested by the offspring.
So, placental mammals can develop more rapidly than marsupials. Some placentals
produce offspring that can walk immediately; these are precocial young. Other
species, like cats and humans, produce offspring that still need considerable
parental care. These offspring are called altricial.
The
Synapsids and the Mammals: The first group to dominate
were the synapsids. They dominated during the Permian, and were represented
by the great "sail-finned" Pelycosaurs and the Therapsid lineages
(the Gorgonopsians, the Dicynodonts, and the Cynodonts). The synapsid lineage
that is alive today are the Mammals. The transition from ancestral to derive
synapsids is well preserved in the fossil record, documenting the evolution
of the inner ear from the ancestral jaw bones, and the evolution of complex
and specialized dentition. The first true mammals are the Morganucodonts that
evolved in the Jurassic Period of the Mesozoic, about 200 mya. Ancestral synapsids
were large carnivores that dominated the landscape; a niche that would be filled
by the diapsid dinosaurs during the Mesozoic. The mammals were small, noctural
insectivores and scavengers. They radiated into these niches in the Jurassic,
perhaps because their endothermy allowed them to use the cool night more effectively
and under cover of darkness from larger predators. The surviving lineage of
this radiation are the three species of monotremes alive today: the platypus
and two species of echidna. These mammals lay eggs. Their offspring hatch, and
then nuzzle the belly of their mothers. Their mothers have sweat glands that
have evolved to produce a nutrient rich milk - they are mammary glands. However,
even the modern representatives lack nipples on these glands - so they "sweat"
milk onto their belly and the the hatchlings lick it up. Placental and marsupial
mammals evolved later, in the Cretaceous Period. Marsupials do not lay eggs;
they give live birth to a very small, embryonic newborn. The newborn climbs
from the genital opening to the pouch - a flap of skin covering the nipples
of the mammary glands. The embryo attaches to a nipple and feeds nearly continuously,
completing development in the pouch. This allows the mother to carry the embryo
with her for a longer period; there are no eggs that must be left periodically
for the mother to feed. The last group of mammals, the placentals, also retain
the embryo and give live birth. However, they retain the embryo much longer,
often until the offspring is capable of independent activity. The efficient
feeding of the embryo is facilitated by the placenta - which allows nutrients
in the bloodstream of the mother to pass directly into the bloodstream of the
offspring. This is far more efficient, energetically, than the mother turning
the nutrients into milk, that must be consumed and digested by the offspring.
So, placental mammals can develop more rapidly than marsupials. Some placentals
produce offspring that can walk immediately; these are precocial young. Other
species, like cats and humans, produce offspring that still need considerable
parental care. These offspring are called altricial.
The Diapsids: The other major group of amniotes are the diapsids. They radiated into two major groups, the lepidosaurs that walked with their legs splayed out and the archosaurs that walked with their legs underneath.
 The
archosaurs would dominate during the Mesozoic Era, with the great
radations of the Crocodylomorphs, Pterosaurs, and Dinosaurs. Two groups of Archosaurs
persists today - the crocodilians (caimen, gavials, alligators, and crocodiles),
and the birds. Although several groups of dinosaurs evolved feathers - probably
first for attracting mates or communicating, and then for insulation - only
in the birds did these feathers evolve for powered flight. Associated shared
derived characters are light, hollow, fused bones, the fusion of digits in the
forelimbss, a keeled sternum for anchoring large breast muscles used for flight,
and a 'wishbone' - a union of the two clavicles that acts like a spring during
the flight stroke. The loss of teeth is also a derived trait in this group,
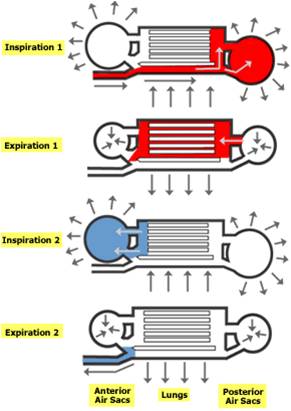
as is the evolution of an interesting respiratory system of air sacs. Birds
have a one way lung. Air that's inhaled goes to a posterior air sac. On the
first exhalation, air is moved forward through the lung. The second inhalation
refills the posterior air sac, and the next exhalation pulls deoxygenated air
forward into the anterior air sac and out of the animal. This modification,
like a one-way digestive tract, improves the efficiency of the organ. Now, the
air in the lung is always oxygenated; it is not mixed with deoxygenated air
like in the sac-like lungs of other animals. It's not surprizing that birds
would evolve this system, as the metabolic demands of flight are much more significant
than other forms of locomotion.
The
archosaurs would dominate during the Mesozoic Era, with the great
radations of the Crocodylomorphs, Pterosaurs, and Dinosaurs. Two groups of Archosaurs
persists today - the crocodilians (caimen, gavials, alligators, and crocodiles),
and the birds. Although several groups of dinosaurs evolved feathers - probably
first for attracting mates or communicating, and then for insulation - only
in the birds did these feathers evolve for powered flight. Associated shared
derived characters are light, hollow, fused bones, the fusion of digits in the
forelimbss, a keeled sternum for anchoring large breast muscles used for flight,
and a 'wishbone' - a union of the two clavicles that acts like a spring during
the flight stroke. The loss of teeth is also a derived trait in this group,
as is the evolution of an interesting respiratory system of air sacs. Birds
have a one way lung. Air that's inhaled goes to a posterior air sac. On the
first exhalation, air is moved forward through the lung. The second inhalation
refills the posterior air sac, and the next exhalation pulls deoxygenated air
forward into the anterior air sac and out of the animal. This modification,
like a one-way digestive tract, improves the efficiency of the organ. Now, the
air in the lung is always oxygenated; it is not mixed with deoxygenated air
like in the sac-like lungs of other animals. It's not surprizing that birds
would evolve this system, as the metabolic demands of flight are much more significant
than other forms of locomotion.
The Lepidosaurs also radiated during the Mesozoic, but did not do so as dramatically as the Archosaurs. Today, the surviving representatives are the Squamata (lizards and the derived snakes) and their sister group, the Sphenodonts with two surviving relict species of Tuatara native to New Zealand. The tuatara maintains the most primitive amniote characteristics, and so is very valuable in studying the phylogeny of diapsid reptiles. The derived squamates, the modern lizards and snakes, have scales, a double hinged jaw (particularly important in snakes), and a hinge on the top of their skull.
That concludes a look at the phyla of animals. Next we will take a brief look at the evolutionary history of our own small twig on the mammalian limb.
Study Questions:
1. What were the probable benefits of colonizing land?
2. What is an amniotic egg and what benefit does it provide? Why did amniotes radiate in the Permian?
3. Describe the reproductive differences among the monotremes, marsupials, and placentals. What advantages were there for marsupial and then placental reproductive strategies?
4. Name the two major groups of diapsids, and the living representatives of each.
5. Explain how a bird's respiratory system is more efficient than a mammal's.
6. What distinguishes the squamates as a group?