 The
Diversity of Life The
Diversity of Life
The
Diversity of Life The
Diversity of Life 1. Overview:
The other major opisthokont group is the Animalia. Animals are distinguished as multicellular, heterotrophic eukaryotes that lack a cell wall. Obviously, the lack of a cell wall distinguishes them from the other multicelluar, heterotropic eukaryotes - the fungi. There is genetic and morphological evidence that animals evolved from choanoflagellate ancestors - probably between 900-700 mya. The oldest animal fossils unearthed to date were found in Australia, in August 2010. They are thought to be sponges, and they date to 650 mya. Sponges are the most 'primitive' group of animals, with the most direct ties to the choanoflagellates, so their early appearance in the fossil record corroborates phylogenetic hypotheses.
 Animal
fossils become much more abundant in the fossil record 550 mya. Although there
are older fossils of soft -bodied jellyfish and worms, the fossil record seems
to 'explode' about 550 mya - an observation that led early Paleotologists to
demarcate this as the beginning of the Paleozoic Era. This explosion probably
occurred because some animals - notably arthropods and molluscs - evolved a
protective body covering (exoskeleton or shell, respectively) that protected
them against the stinging cnidarian predators. The radiation of these groups
left a disproportionate mark on the fossil record because hard parts fossilize
much more readily than soft tissue. So, in part, the 'Cambrian Explosion' is
probably due to sampling bias - organisms evolved that were more likely to fossilize,
so we see more fossils. The evolution of arthropods in the Cambrian led to an
explosion of diversity that is still occurring today - over 85% of the animal
species named to date are Arthropods. In the figure to the right, we see an
artists depiction of the Cambrian Period, with a variety of sponges, cnidarians,
molluscs, and arthropods, including the large predatory arthropod, Anomalocaris
- that grasped other animals with its raptorial claws and crushed them in their
circular jaws.
Animal
fossils become much more abundant in the fossil record 550 mya. Although there
are older fossils of soft -bodied jellyfish and worms, the fossil record seems
to 'explode' about 550 mya - an observation that led early Paleotologists to
demarcate this as the beginning of the Paleozoic Era. This explosion probably
occurred because some animals - notably arthropods and molluscs - evolved a
protective body covering (exoskeleton or shell, respectively) that protected
them against the stinging cnidarian predators. The radiation of these groups
left a disproportionate mark on the fossil record because hard parts fossilize
much more readily than soft tissue. So, in part, the 'Cambrian Explosion' is
probably due to sampling bias - organisms evolved that were more likely to fossilize,
so we see more fossils. The evolution of arthropods in the Cambrian led to an
explosion of diversity that is still occurring today - over 85% of the animal
species named to date are Arthropods. In the figure to the right, we see an
artists depiction of the Cambrian Period, with a variety of sponges, cnidarians,
molluscs, and arthropods, including the large predatory arthropod, Anomalocaris
- that grasped other animals with its raptorial claws and crushed them in their
circular jaws.
Animals are a fairly recent addition to the living world. They are not very genetically or metabolically diverse when compared to prokaryotes or protists that have much longer evolutionary histories. However, animals are structurally diverse. They have radiated into a wide variety of shapes through changes in developmental patterning. These different shapes have allowed them to exploit a wide range of habitats and resources. Animals are classified into phyla based on their shape or body plan. As heterotrophs, these differences often correlate with differences in feeding structures or locomotory structures that they use to capture food.
From our vantage point as large terrestrial organisms, we have a very skewed view of animal diversity. This is represented by our casual division of animals into 'vertebrates' (like us) and 'invertebrates' (other stuff not like us). These are not phylogenetic groups. The Vertebrata is one sub-phylum of the phylum Chordata, which is one of 32 phyla of animals. The two other sub-phyla of Chordates, and all the other 31 phyla, are 'invertebrate' - and they obviously represent the bulk of animal diversity. Indeed, 95% of all animal species are 'invertebrates', with 85%, as mentioned, occurring in the single phylum Arthropoda. In addition, most animal phyla are exclusively marine - so we rarely encounter them. As you look through the groups pictured at the top of the page (the protostome phyla), you'll see that most animals are rather simple, bilaterally symmetrical, worm-like organisms with different feeding structures.
We are only going to consider the major phyla of animals here, those representing important evolutionary innovations in the group, or representing important radiations.
2. Development
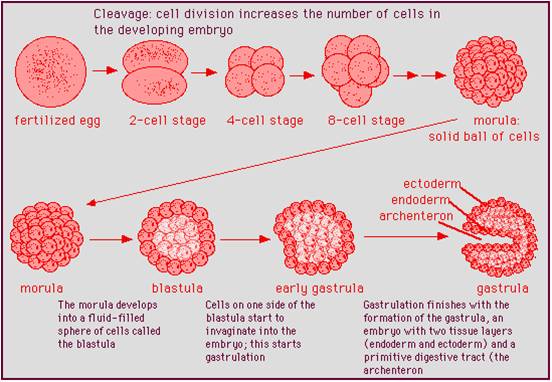 One
of the defining features in the animals is their method of early embryonic development.
The subsequent differences that develop in shape occur after these initial stages
take place. The union of sperm and egg form a diploid zygote. the zygote divides
by mitosis, forming a solid ball of cells called the morula. The cells in the
morula migrate, moving out from under other cells until they form a hollow ball
of cells called the blastula. Then, one side of the blastula migrates in towards
the center of the embryo. This process of infolding, or invagination, creates
the gastrula. The gastrula has two layers of cells: one layer is on the outside
(ectoderm) and the other (endoderm) is on the inside, lining the newly-formed
cavity the infolding produced. This cavity becomes the gut in all animals. The
hole that connects the gut with the environment becomes either the mouth or
anus of the mature animal. Although sponge development is slight different,
they still form these typical stages. In all other animals, from cnidarians
to cats, the process is fundamentally the same. Differences in shape emerge
as the embryo developes from this gastrula.
One
of the defining features in the animals is their method of early embryonic development.
The subsequent differences that develop in shape occur after these initial stages
take place. The union of sperm and egg form a diploid zygote. the zygote divides
by mitosis, forming a solid ball of cells called the morula. The cells in the
morula migrate, moving out from under other cells until they form a hollow ball
of cells called the blastula. Then, one side of the blastula migrates in towards
the center of the embryo. This process of infolding, or invagination, creates
the gastrula. The gastrula has two layers of cells: one layer is on the outside
(ectoderm) and the other (endoderm) is on the inside, lining the newly-formed
cavity the infolding produced. This cavity becomes the gut in all animals. The
hole that connects the gut with the environment becomes either the mouth or
anus of the mature animal. Although sponge development is slight different,
they still form these typical stages. In all other animals, from cnidarians
to cats, the process is fundamentally the same. Differences in shape emerge
as the embryo developes from this gastrula.
3. Body Plans
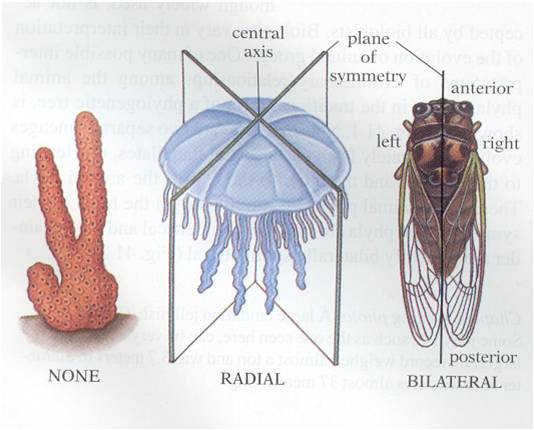 Animals
exibit a wide range of shapes, but these shapes can be classified in three major
groups. First, the sponges are asymmetrical. Second, the cnidarians and ctenophores
are 'radially symmetrical. This means that, if you are looking down on the organism
from the top, you could cut the organism is half along any axis passing through
the midpoint and the two halves would be be the same. The rest of the animals
are bilaterally symmetrical. This means that there is only one plane of division
that will create to symmetrical 'sides' (bi-lateral = two sides). Given the
fact that the vast majority of animal phyla are bilaterally symmetrical, we
can hypothesize that this evolutionary innovation was functionally important
- allowing animals to work "better", somehow. But how?
Animals
exibit a wide range of shapes, but these shapes can be classified in three major
groups. First, the sponges are asymmetrical. Second, the cnidarians and ctenophores
are 'radially symmetrical. This means that, if you are looking down on the organism
from the top, you could cut the organism is half along any axis passing through
the midpoint and the two halves would be be the same. The rest of the animals
are bilaterally symmetrical. This means that there is only one plane of division
that will create to symmetrical 'sides' (bi-lateral = two sides). Given the
fact that the vast majority of animal phyla are bilaterally symmetrical, we
can hypothesize that this evolutionary innovation was functionally important
- allowing animals to work "better", somehow. But how?
Bilateral symmetry does more than create two sides; it also means that there is a top and bottom, and a front and back. As heterotrophs, most animals have to moving through the environment in search for their food. Having a 'front' would favor the concentration of sensory systems at this end - so that the animal could perceive the environment that it was about to enter and search for food and avoid predators. So, bilateral symmetry is associated with the formation of a head and the evolution of a brain for processing all this sensory information and directing locomotion. It is no coincidence, therefore, that bilateral symmetry allowed some animals to "get a-head". hahaha.
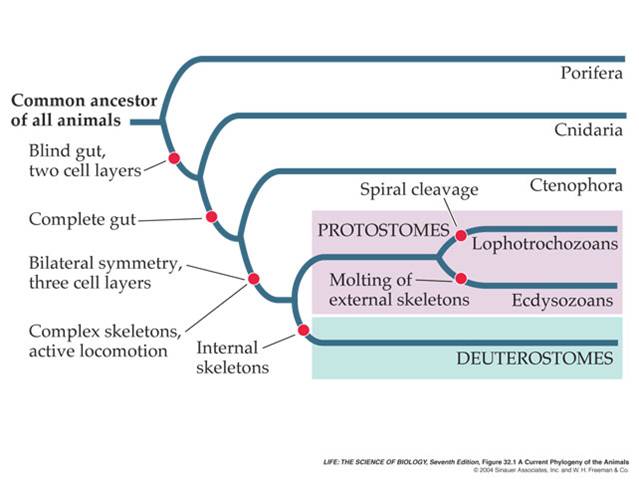 4.
Phylogeny
4.
Phylogeny
These developmental patterns are reflected by animal phylogeny. Sponges are the simplest animals; they retain the collar cells (choanocytes) that are so similar to the choanoflagellates, and the cells in sponges do not form true tissue layers (where the cells are bound together in a matrix of secreted proteins). The remaining animals are distinguished by having only two true tissues (ecto and endoderm) or three (with mesoderm). In addition, the more primitive animals are radially symmetrical, while the animals with three tissue layers are bilaterally symmetrical. These bilateral animals are grouped by whether the blastopore forms the mouth ('protostomes') or anus ('deuterostomes'). The protostomes are further classified by how their larvae develop or how they grow: lophotrochozoans have particualr larval stages and grow by adding segments in some cases, whereas the ecdysozoans have a cuticle/exoskeleton and must shed this in order to grow. We will review these patterns as we move through the major animal phyla.
5. Major Phyla
a. Phylum Porifera: Sponges
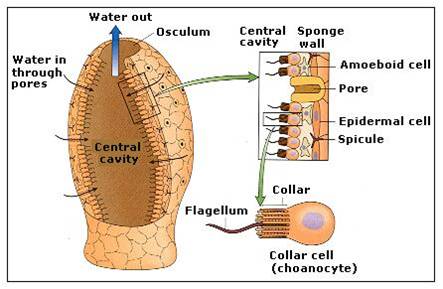 Sponges
consist of specialized but loosely integrated cells; the cells are not
anchored in a layer of protein (known as a basement membrane) as in
other animals. So, curiously, some sponges can be 'disintegrated' in a blender
and will then re-assemble into many small sponges, with cell types reintegrating
into their appropriate relationship. These aggregates are asymmetrical, but
the fundamental filtering unit is often vase-like. The cells in a vase unit
form two layers; the inside layer consists of choanocytes that are very similar
to free-living choanoflagellates. The beating of the flagella create a current
that pulls water through pore cells into the central chamber of the sponge -
water exits through the top hole or 'osculum'. The choanocytes feed by ingesting
organic material by phagocytosis. The outer cell layer is the 'epidermis'. The
epidermis receives nutrients from the choanocytes in an interesting way: the
food that is absorbed by the inner layer of choanocytes is passed to amoebocytes
- motile cells that live in the space between the cell layers. This space, the
mesohyl, contains flexible spongin proteins, and may contain rigid spicules
of silca. The ameobocyte crawls back and forth, transferring food from the chaoanocytes
to the epidermal cells. Sponges are filter feeders. They reproduce asexually
by fragmentation (pieces can break off and settle elsewhere), and by sexual
reproduction.
Sponges
consist of specialized but loosely integrated cells; the cells are not
anchored in a layer of protein (known as a basement membrane) as in
other animals. So, curiously, some sponges can be 'disintegrated' in a blender
and will then re-assemble into many small sponges, with cell types reintegrating
into their appropriate relationship. These aggregates are asymmetrical, but
the fundamental filtering unit is often vase-like. The cells in a vase unit
form two layers; the inside layer consists of choanocytes that are very similar
to free-living choanoflagellates. The beating of the flagella create a current
that pulls water through pore cells into the central chamber of the sponge -
water exits through the top hole or 'osculum'. The choanocytes feed by ingesting
organic material by phagocytosis. The outer cell layer is the 'epidermis'. The
epidermis receives nutrients from the choanocytes in an interesting way: the
food that is absorbed by the inner layer of choanocytes is passed to amoebocytes
- motile cells that live in the space between the cell layers. This space, the
mesohyl, contains flexible spongin proteins, and may contain rigid spicules
of silca. The ameobocyte crawls back and forth, transferring food from the chaoanocytes
to the epidermal cells. Sponges are filter feeders. They reproduce asexually
by fragmentation (pieces can break off and settle elsewhere), and by sexual
reproduction.
b. Phylum Cnidaria: Hydra, anemones, corals, jellyfish
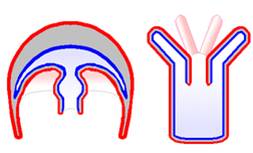 Although
the cnidarians may seem to be quite different from one another, the body plans
of jellyfish ('medusa' plan) and anemones ('polyp' plan) are actually very similar.
All cnidarians have a rather simple plan of two true tissue layers separated
by a gel/protein filled space (mesoglea). The outer layer covers the tentacles,
which have cnidocytes. These are cells that house a harpoon-like structure.
Any bump or brush and the harpoon is deployed, along with toxins that paralyze
the prey. The tentacles contract and draw the prey into the gut cavity, where
extracellular digestion occurs. Emzymes are produced by endodermal tissue and
these enzymes are secreted into the gut cavity. The products of this digestion
are absorbed by cells by phagocytosis, so there is both intra and extracellular
digestion. As in sponges, there are ameobocytes that transfer nutrients from
the endodermis to the ectodermis. The inner cell layer, the endoderm, lines
the gut cavity. In cnidarians, digestion is both extracellular and intracellular.
Cnidarians have a 'nerve net' - a decentralized organization of nerve-like cells
that coordinate the movement of tentacles. Some cnidarians are exclusively asexual,
but most can reproduce sexually.
Although
the cnidarians may seem to be quite different from one another, the body plans
of jellyfish ('medusa' plan) and anemones ('polyp' plan) are actually very similar.
All cnidarians have a rather simple plan of two true tissue layers separated
by a gel/protein filled space (mesoglea). The outer layer covers the tentacles,
which have cnidocytes. These are cells that house a harpoon-like structure.
Any bump or brush and the harpoon is deployed, along with toxins that paralyze
the prey. The tentacles contract and draw the prey into the gut cavity, where
extracellular digestion occurs. Emzymes are produced by endodermal tissue and
these enzymes are secreted into the gut cavity. The products of this digestion
are absorbed by cells by phagocytosis, so there is both intra and extracellular
digestion. As in sponges, there are ameobocytes that transfer nutrients from
the endodermis to the ectodermis. The inner cell layer, the endoderm, lines
the gut cavity. In cnidarians, digestion is both extracellular and intracellular.
Cnidarians have a 'nerve net' - a decentralized organization of nerve-like cells
that coordinate the movement of tentacles. Some cnidarians are exclusively asexual,
but most can reproduce sexually.
The "Bilateria"
The remaining animal phyla are bilaterally symmetrical organisms. Their symmetry is encoded by Hox genes that establish the anterior-posterior axis of the body. As mentioned above, bilateral symmetry is advantageous because it establishes a polarity to the animal - and sense systems and neaural integration can be concentrated at the anterior end.
The bilaterally symmetrical animals have three true tissue layers (triploblastic), adding mesoderm to endo- and ectoderm. Tehy form two major clades, distinguished by whether the blastopore developes into the mouth (Protostomes) or the anus (Deuterostomes) in the adult. The protostomes are further divided by other developmental characteristics. One major clade, the Ecdysozoa, must shed a hard cuticle or exoskeleton in order to grow. The other clade, the Lophotrochozoans, is distinguished by having either a rake-like feeding apparatus ('lophophore' - seen in some of the pictures in the lower two rows, above), or a specific larval stage known as a 'trochophore'. As you can see from the phylogeny above, there are many groups whose phylogenic relationships are unknown at this time.
Study Questions:
1. What are the stages of animal embryogenesis?
2. Distinguish "protostome" and "deuterostome".
3. Why are "goofy" characteristics like whether the blastopore forms the mouth or anus used to reconstruct phylogenies? Explain in terms of the three reasons why organisms might share a characteristic.
4. Draw the cell layers in a sponge and describe the function of each.
5. What are the two body types seen in Cnidarians?
6. List four groups of Cnidarians.
7. How do Cnidarians feed?
8. Why is bilateral symmetry, rather than radial symmetry, associated with "cephalization" in motile organisms?
9. Draw the phylogeny of animals, including the characteristics of each major branch.