(a
Homopteran... a true bug) - A single sex chromosome - 6 autosomal, no sex chromosome
(female)
6 autosomes, plus sex chromosome (male)
b.
Lygaeus (another bug) - two sex chromosomes:
- In humans and most mammals, the heterogametic sex is male (XY) and the homogametic
sex if female (XX). In humans and most mammals, the presence of a "y" chromosome
causes male development, even if the individual has 2 X's by mistake (XXY).
At least, usually individuals that are XY are male. Actually, 'maleness'
is determined by only a small region of the Y - called the "sex determining
region of the y" - SRY - gene. This single gene can be lost from the chromosome
through mutation, and sometimes it can even be transferred to the X during male
sperm production. So, this man produces "y" sperm that LACK the sry, and "X"
sperm that have the sry. (This only occurs in the couple sperm where this mutation
occurred; it would not occur in all this man's sperm. So, this is VERY rare.
This sry "X" chromosome, when paired with the X of the mother, would produce
a zygote that is XX but has the sry and would develop into a male child. Likewise,
if a sperm with the mutant Y is used, then a zygote is produced that is XY but
has no sry gene and developes into a female.
- In birds, some reptiles, and butterflies
and moths, the homogametic sex is male (WW) whereas the heterogametic sex (ZW)
is female. We use the Z W symbols to represent sexes in these species.
B. Gender
 Gender
is a role or behavior that a society correlates with a sex. These 'gender-roles'
and behaviors vary across societies, even though most humans are biologically
male or female. So, in the USA, we might associate the wearing of make-up and
dressing in skirts as playing the gender role of a woman. In the Wodaabe tribe
of the Sahel region of Africa, these cultural practices are associated with
the gender-role of a man. And don't dare tell a scotsman he is wearing a skirt...
it's a kilt! In some societies, the expected 'social norm' of sexual preference
for someone playing the gender role of a man is to be attracted to women. In
ancient Greece, sexual behavior was not as strongly correlated with our gender
categories of 'man' and 'woman', but by the categories of the 'active role'
(adult, penetrative, maculine, higher social status) and 'passive role' (youth,
feminine, lower social status). These roles could be played by members of the
same or opposite sex.
Gender
is a role or behavior that a society correlates with a sex. These 'gender-roles'
and behaviors vary across societies, even though most humans are biologically
male or female. So, in the USA, we might associate the wearing of make-up and
dressing in skirts as playing the gender role of a woman. In the Wodaabe tribe
of the Sahel region of Africa, these cultural practices are associated with
the gender-role of a man. And don't dare tell a scotsman he is wearing a skirt...
it's a kilt! In some societies, the expected 'social norm' of sexual preference
for someone playing the gender role of a man is to be attracted to women. In
ancient Greece, sexual behavior was not as strongly correlated with our gender
categories of 'man' and 'woman', but by the categories of the 'active role'
(adult, penetrative, maculine, higher social status) and 'passive role' (youth,
feminine, lower social status). These roles could be played by members of the
same or opposite sex.
 In
almost all social animals, there are correlations between sex and social behavior.
But, as in humans, these behaviors can vary between populations and between
closely related species. Bonobo chimps, for instance, mate face-to-face like
humans, and are different from common chimps to whom they are more closely related.
Indeed, sex is used as a form of communication in bonobos - it has become a
ritualized behavior of communication within and between sexes, often used as
a greeting between any two bonobos, male or female. Many other species also
engage in frequent sexual behaviors between members of the same sex; 90% of
observed sexual pairings in giraffes occurs between males. So, sex in giraffes
may be like sex in Ancient Athens - a way to communicate dominance. Or, as in
bonobo's, it may just be done for fun and social bonding. There are lots of
cases where organisms use sex for reasons other than pro-creation. And, in almost
all social species, there is a fraction of individuals that choose same-sex
pairings, or perform the behavioral gender roles typical for the other sex.
In some species, individuals even use gender roles to deceive others in their
own population. For example, in cuttlefish (relatives of squid) young males
color themselves as females and enter the territory of dominant males. They
play the gender role of a female, and are allowed into the territory. When the
dominant male isn't looking, the immature male serruptitiously mates with females
in the territory. These males are called 'sneaker' males. The same thing happens
in many birds, where young males maintain their immature, female-like plumage
and gain access to male territories where they mate with females. As with nearly
everything we've examine in biology, a more rigorous examination of patterns
in nature tend to reveal continua; even between categories that, at the outset,
seem discrete: categories like species, sexes, gender, and sexual behavior.
In
almost all social animals, there are correlations between sex and social behavior.
But, as in humans, these behaviors can vary between populations and between
closely related species. Bonobo chimps, for instance, mate face-to-face like
humans, and are different from common chimps to whom they are more closely related.
Indeed, sex is used as a form of communication in bonobos - it has become a
ritualized behavior of communication within and between sexes, often used as
a greeting between any two bonobos, male or female. Many other species also
engage in frequent sexual behaviors between members of the same sex; 90% of
observed sexual pairings in giraffes occurs between males. So, sex in giraffes
may be like sex in Ancient Athens - a way to communicate dominance. Or, as in
bonobo's, it may just be done for fun and social bonding. There are lots of
cases where organisms use sex for reasons other than pro-creation. And, in almost
all social species, there is a fraction of individuals that choose same-sex
pairings, or perform the behavioral gender roles typical for the other sex.
In some species, individuals even use gender roles to deceive others in their
own population. For example, in cuttlefish (relatives of squid) young males
color themselves as females and enter the territory of dominant males. They
play the gender role of a female, and are allowed into the territory. When the
dominant male isn't looking, the immature male serruptitiously mates with females
in the territory. These males are called 'sneaker' males. The same thing happens
in many birds, where young males maintain their immature, female-like plumage
and gain access to male territories where they mate with females. As with nearly
everything we've examine in biology, a more rigorous examination of patterns
in nature tend to reveal continua; even between categories that, at the outset,
seem discrete: categories like species, sexes, gender, and sexual behavior.
C. Sex Linkage
Sex linkage describes the pattern
of heredity between traits governed by genes on the sex chromosomes and the
trait of 'sex', itself. In humans, although the Y is a small chromosome with
little besides the sry gene, the X is a large chromosome with hundreds of genes
that govern a wide variety of phenotypic traits. Since the X chromosome influences
sex, and also carries genes that influence these traits, the pattern of heredity
of sex and these X-linked genes is not independent; whereever the X goes (and
influences sex), these other genes go, too. So,
the pattern of heredity of these genes is NOT independent of sex - violating
the pattern of independent assortment between sex and these x-linked traits.
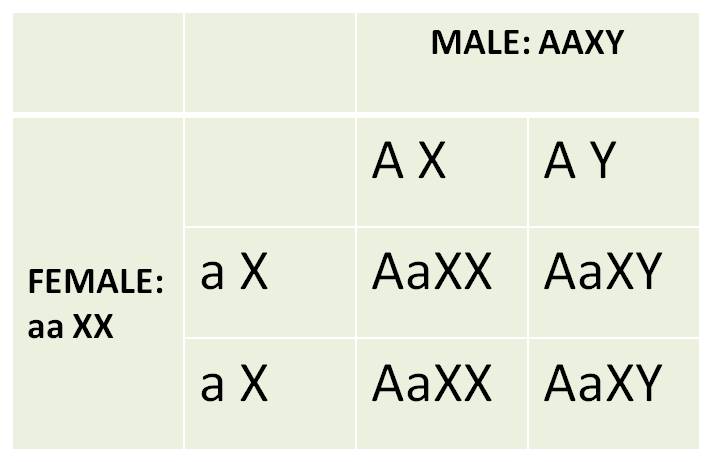
1. For Comparision:
The Independent Assortment of Sex (as a trait) and an Autosomal Trait:
Genes that are not on sex chromosomes
are called autosomal genes. So, suppose we followed the pattern of heredity
of "tongue rolling" and sex in humans. Tongue rolling is the ability
to curl your tongue into a tube, by lifting the edges of your tongue up. It
is a trait that is dominant ('A') to the inability to roll the tongue ('a').
Because this trait is autosomal, it is governed by genes on a pair of homologous
chromosomes that are NOT the sex chromsomes. So, the way the autosomes pair
during gamete formation will be independent of how the sex chromosomes pair,
and the genes they carry will be inherited independently of one another. So,
suppose we do reciprocal dihybrid crosses:
 |
 |
100%
tongue rolling in sons and daughters |
100%
tongue rolling in sons and daughters |
As expected, the results of the reciprocal
crosses are the same, and are consistent with Mendel's predictions for independently
assorting traits; the sex of the tongue-rolling parent doesn't matter - results
in the reciprocal crosses are the same. Now, let's see what happens when we
consider two traits governed by genes on the same chromosomes - in this case,
with sex-linked genes, one trait is "sex" and the other trait is colorblindness.
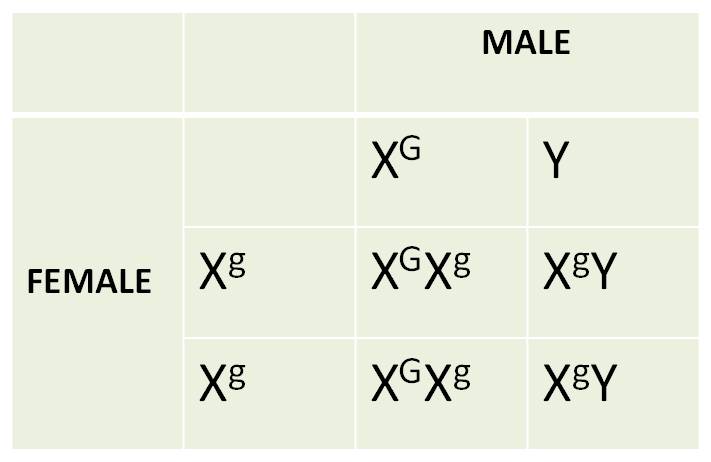
2. An Example
of Sex-Linkage: Red-Green Colorblindness in Humans:
There are many forms of 'color blindness'.
Two of these are caused by alleles on the X chromosome. In one case, the red-detecting
photoreceptors are absent, and a person sees red as black. In another form,
the green photoreceptors are absent, affect the ability to distinguish red from
green. As with all X linked traits, these conditions are expressed in males
more frequently than in females. When males inherit an X with these alleles,
they must express the trait; they have no other chromosome that carries these
genes, because as males, their other sex chromsomes is a Y that does not carry
these genes. So, consider this set of reciprocal crosses:
 |
 |
100%
normal vision |
50%
colorblind (all males): 50% normal (all females) |
The results of the reciprocal cross
are DIFFERENT. Unlike Mendel's experiments where the sex of the parent giving
a trait didn't matter, here it does; sons look like their mothers. Even when
the mother does not express the trait, but carries a recessive allele in the
heterozygous condition, there is a 50% chance that her son will inherit the
allele on their X and expressing the trait. Males with normal vision will only
have daughters with normal vision, because they will pass their one normal X
to all their daughters.
Heredity, Gene Regulation, and Development
I. Mendel's Contributions
II. Meiosis and
the Chromosomal Theory
III. Allelic, Genic,
and Environmental Interactions
IV. Sex Determination
and Sex Linkage
V. Linkage
A. Overview
Sex linkage showed us that traits
can be inherited in a related manner, not just independently. In sex linkage,
the transmission and expression of a trait correlated with the transmission
of sex chromosomes, so there was a relationship between expressing that trait
and expressing a particular sex. Curiously, for x-linked human traits, the relationship
is indirect. The gene that causes maleness (sry) is on the Y, but most sex-linked
genes are on the X; so the correlation with sex is due to the combination
of sex chromosomes received, not due to the inheritance of a single
chromosome that governs BOTH these traits.
The pattern with which pairs (or
sets) of genes are inherited can actually be much more direct. For example,
suppose we consider the pattern of heredity for traits governed by genes on
the same chromosome? Then obviously, their pattern of heredity will be directly
related. It is this scenario that we examine in this lecture - the pattern of
heredity of genes that are linked together as part of the same chromosome.
A critical aspect of this lecture
will be understanding the process of crossing-over. As you recall, as the chromosomes
condense in Prophase I of meiosis, they condense with their homolog. During
this condensation period, homologs can exchange pieces of chromosome. This is
called crossing over. Now, because these are homologous chromosomes that have
the same loci, an equal exchange means that both chromosomes still have a gene
for every trait they govern. However, because the pieces they exchange may contain
different alleles for these genes, crossing over can produce new combinations
of alleles along the length of each chromosome that did not exist before. You'll
see how this works as we go along.
There are some assumptions that we
will make here about crossing over:
1) Crossing over is a random event.
Where a cross-over occurs along a chromosome is random. (Geneticists have found
that this is not explicitly true - there are places where crossing over is more
likely to occur...but we will make this assumption here.)
2) So, the greater the DISTANCE between
genes on a chromosome, the more likely it is that a random cross-over event
will occur between them. Likewise, the probability of a cross-over event declines
as the distance between genes declines.
3) So, for genes in a local region
of a chromosome, crossing over will be a rare event.
4) And, given this relationship,
it means that we can use the frequency of crossing over, if we can measure it,
as an index of the distance between the genes. YOU NEED TO UNDERSTAND THIS LOGIC
BEFORE GOING ANY FURTHER.
B. Complete Linkage
Let's start with the most extreme
(and simplest) example. Consider two loci, A and B, that are immediate
neighbors on a chromosome; so close that they are always inherited together
because the chance that a random cross-over event splits them exactly is ridiculously
small.
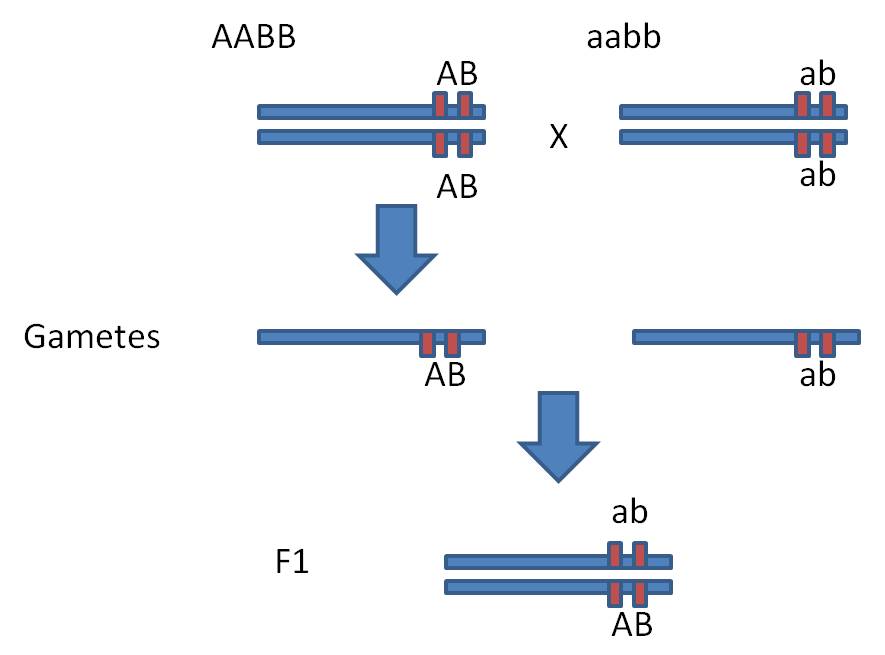 Suppose
we do the following dihybrid cross:
Suppose
we do the following dihybrid cross:
Dihybrid Cross: AABB x aabb = 100% AaBb.
Hmmm... well, so far the results
are indistinguishable from independently assorting genes.
 But
now do the F1 x F1 cross. Each parent can only produce two types of gametes,
because the AB genes and the ab genes are parts of the same chromosomes and
do not assort independently. In this case, wherever the "A" goes,
the "B" has to go, too; they are part of the same unit. GIVEN THIS
spatial relationship (with AB linked and ab linked) and no crossing over, each
parent can only produce AB and ab gametes types (and not the additional Ab and
aB types we would expect from an AaBb parent with independent assortment).
[*Of course, the other spatial arrangment
is possible, with the Ab linked together and the aB linked together. But we
would have had to start with different pure-breeding stocks in the parental
generation (AAbb, aaBB). And, if this was the arrangment, then the F1 would
only be able to produce Ab and aB gametes. OK, back to the problem we have!]
But
now do the F1 x F1 cross. Each parent can only produce two types of gametes,
because the AB genes and the ab genes are parts of the same chromosomes and
do not assort independently. In this case, wherever the "A" goes,
the "B" has to go, too; they are part of the same unit. GIVEN THIS
spatial relationship (with AB linked and ab linked) and no crossing over, each
parent can only produce AB and ab gametes types (and not the additional Ab and
aB types we would expect from an AaBb parent with independent assortment).
[*Of course, the other spatial arrangment
is possible, with the Ab linked together and the aB linked together. But we
would have had to start with different pure-breeding stocks in the parental
generation (AAbb, aaBB). And, if this was the arrangment, then the F1 would
only be able to produce Ab and aB gametes. OK, back to the problem we have!]
So, in the F1 x F1 cross, these two
genes are inherited as if they were one gene; yielding a 3:1 ratio of AB:ab
offspring.
The critical test, of course, is
to see what type of gametes this F1 individual can make. And to determine what
type of gametes an individual can make, what type of cross do we conduct? Correct,
a test cross. In this case, the Test Cross would be: AaBb x aabb. If AB and
ab are completely linked, as above, then the progeny would be = 50%AB (AaBb)
and 50% ab (aabb); with no Ab or aB offspring because this F1 individual would
not make those gamete types.
C. Incomplete Linkage
OK, hopefully you have the idea.
Now let's consider the case where we are studying a pair of genes that are in
the same region of a chromosome, so that some (rare) crossing over will occur
between them. Our goals are:
1) Determine if the genes are linked,
or are assorting independently.
2) If they are linked, detemine the
arrangement of alleles in the F1 individual; which alleles are paired on each
homolog?
3) Determine the distance between
loci.
1. Determining
if the genes are linked, or are assorting independently:
To do this, we will conduct a Test Cross, so that we can "see" the
types of gametes that our F1 individual makes. Then, we can compare these frequencies
with expectations derived from the product rule for independent events.
Suppose you did your Dihybrid test
cross and got: 43 AB,12 Ab, 8 aB, 37ab
Could these genes be assorting independently?
Well, anything is possible, but ARE THESE RESULTS LIKELY from independently
assorting genes? How likely? Let's conduct a Chi-Square test of Independence
to determine the likelihood of observing these results from independent events.
Determine what you would expect
if the genes assorted independently:
- For the A trait, by itself, there are 55/100 A and 45/100 a.
- For the B trait, by itself, there are 51/100 B and 49/100 b.
- So, if the genes assort independently, you can predict the probability of
phenotypic combinations by multiplying the independent probabilities together.
- Now, multiplying fractions
gives you a fraction. To find the number/frequency of individuals, you have
to multiply this final fraction times the sample size (100 in this case). So,
your expected frequencies, under the hypothesis of independent assortment, are:
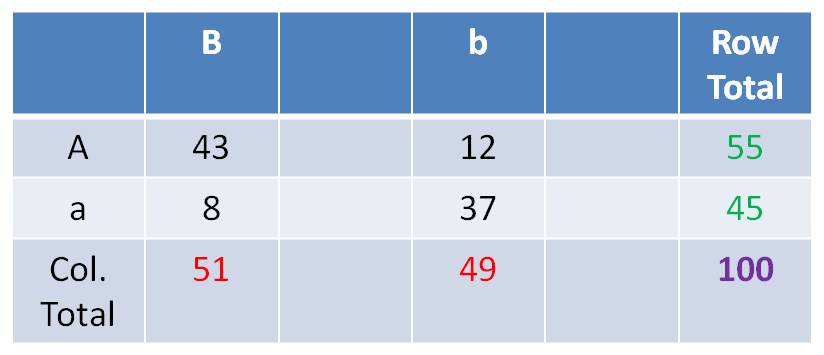 The
frequency of ‘AB’ should = f(A) x f(B) x N = 55/100 x 51/100 x 100
= 28
The
frequency of ‘AB’ should = f(A) x f(B) x N = 55/100 x 51/100 x 100
= 28
The frequency of ‘Ab’ should = f(A) x f(b) x N = 55/100 x 49/100
x 100 = 27
The frequency of ‘aB’ should = f(a) x f(B) x N = 45/100 x 51/100
x 100 = 23
The frequency of ‘ab’ should = f(a) x f(b) x N = 45/100 x 49/100
x 100 = 22
You can calculate these in a 2 x
2 Chi-square Test of Independence Table as:
(Row Total x Column Total)/ Grand Total.
So, the expected frequency of AB
individuals = (51 x 55)/100 = 28
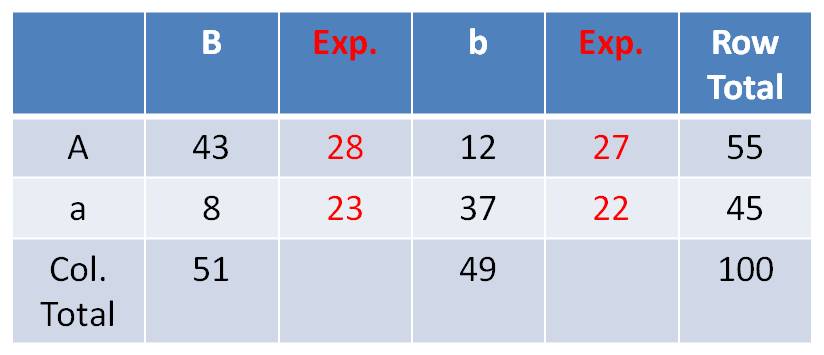 Compare
observed vs. expected using the chi-square computation: X2 = SUM
[(o-e)2/e]:
Compare
observed vs. expected using the chi-square computation: X2 = SUM
[(o-e)2/e]:
| |
Obs. |
Exp. |
(o-e) |
(o-e)2/e |
| AB |
43 |
28 |
15 |
8.04 |
| Ab |
12 |
27 |
-15 |
8.33 |
| aB |
8 |
23 |
-15 |
9.78 |
| ab |
37 |
22 |
15 |
10.23 |
| |
|
|
Sum = |
36.38 |
- Interpreting
the Chi-Square Value of 36.38 - Statisticians have determined how likely particular
chi-square values are, just by chance, if the events truly ARE independent.
They have determined that, with 1 'degree of freedom' (what you have:
(rows - 1)(columns - 1) )), independently occurring events will only produce
a score of 3.84 only 5% of the time. A score of 6.63 will only occur 1% of the
time, just by chance, when two events truly ARE independent. Larger scores are
progressively less likely, because greater deviations from the expectations
are less probable (for events that ARE independent). Your score of 36.38 signifies
that truly Independently Assorting genes would ONLY yield results this deviant
from what you would expect less than 1% of the time. - So, independently assorting
genes are unlikely to yield your results. There is less than 1 chance in 100
that independently assorting genes would produce results that deviate this much
from expectations. Can I say it any other way?
- So, the most appropriate conclusion
is to reject the hypothesis of independence, and conclude that your genes are
NOT assorting independently. You should conclude that they are linked.
HAVING CONCLUDED THAT THE GENES
ARE LINKED, WE CAN NOW ANSWER TWO OTHER QUESTIONS.
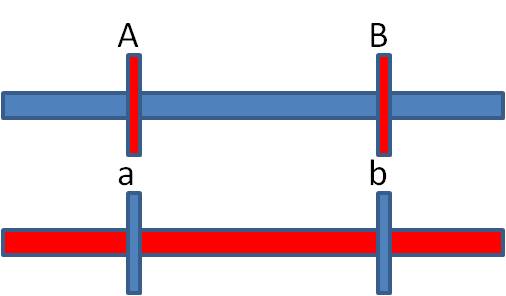 2)
Detemining the arrangement of alleles in the F1 individual; which alleles are
paired on each homolog?
2)
Detemining the arrangement of alleles in the F1 individual; which alleles are
paired on each homolog?
If crossing over is rare, then most
of the time it does NOT occur. So, most of the time, the alleles are passed
to the offspring in their original arrangement in the F1 individual - and these
phenotypes should be the most abundant phenotypes in the offspring.
In our case, these are the AB and ab phenotypes. So, the original AaBb parent
had the A and B alleles on one chromosome and the a and b alleles on the other
homologous chromosome. MOST OF THE TIME, the allelic combinations were inherited
as a unit. These gametes are called the "parental types", because
the preserve the original spatial combination of alleles that existed in the
parent.
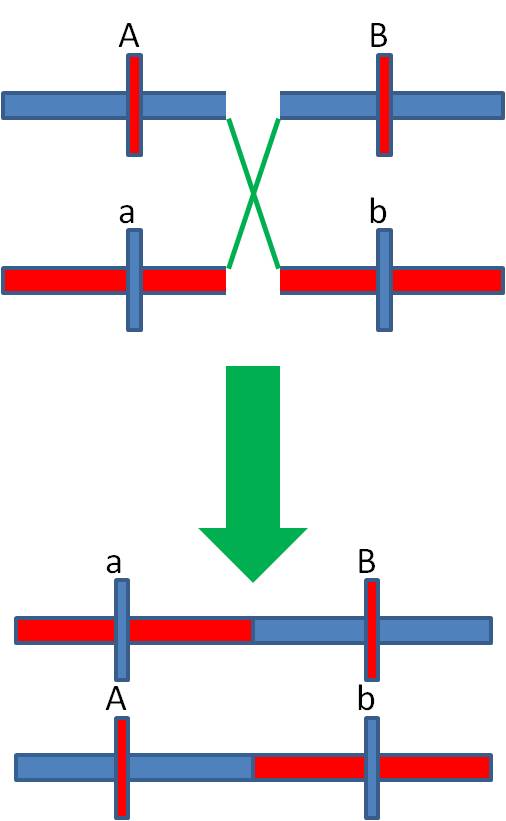 When
a rare cross-over occurs between these loci, the Ab and aB gamete types are
produced. These are less common than the parental types (becasue crossing over
is rare), and they are called the "recombinant types".
When
a rare cross-over occurs between these loci, the Ab and aB gamete types are
produced. These are less common than the parental types (becasue crossing over
is rare), and they are called the "recombinant types".
3. Determining
the distance between loci:
Well, now we apply another assumption:
that the frequency of crossing over is proportional to the distance between
loci. So, we measure the frequency of crossing over and can use that as a direct
index of the distance between loci. So, there were 12 Ab and 8 aB offspring...
for a total of 20 offspring produced by recombination events. This is 20.0%
(20/100 offspring). So, we say that the genes are 20.0 map units (or centiMorgans)
apart. Genes that are closer together will recombine less often... genes that
are farther apart will recombine more often.
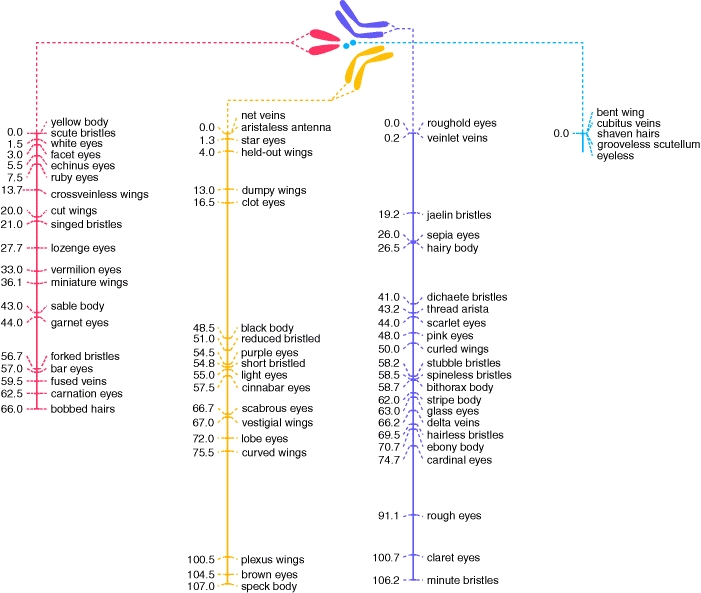 D.
Summary
D.
Summary
So, by observing the frequency with
which pairs of traits are inherited, we can build maps of the physical location
of genes on chromosomes. The pioneers of this method were T. H. Morgan, who
first described sex linkage in the early 1910's, and his student A. Sturtevant,
who conducted many breeding experiments with Drosophila and began construction
of what is known as a linkage map. The one shown here is a partial linkage map
for Drosophila - constructed through heredity experiments using fruit
flies. Linkage maps are very important, and they remain important today, even
though we now have other ways to map DNA - like by the sequence of A, C, T,
and G. However, sequence maps don't show us the spatial relationship of genes
that govern particular traits. In order to know which sequence might code for
a given trait, we need to compare the known location for a sequence with the
known location of a gene governing a trait from a linkage map. This can actually
be very difficult, because the scales of resolution are so different. Through
construction of a linkage map, we might estimate that a trait is governed by
a gene that is 5 centiMorgans from the centromere of the chromosome. But each
centiMorgan covers thousands of base pairs, and the map location of this gene
covers thousands of base pairs, too. Finding the particular sequence within
this region still requires additional work. So, mapping through linkage analysis
is still very important, and we are still finding the locations of human genes
based on the frequency with which they are inherited with known genes, or known
DNA sequences called 'markers'.
Heredity, Gene Regulation, and Development

I. Mendel's Contributions
II. Meiosis and
the Chromosomal Theory
III. Allelic, Genic,
and Environmental Interactions
IV. Sex Determination
and Sex Linkage
V. Linkage
VI. Mutation
A. Overview
Mutation is defined as a change in
the genome. Mutations can occur due to damage to the DNA by mutagens or by errors
in DNA replication. If these changes are not repaired before division, they
are passed to daughter cells. Mutations occuring in body cells are called 'somatic
mutations', and (unless they occur in cells that will become ovaries or testes)
they will not be passed to offspring. However, mutations can also occur in reproductive
tissues - in the cells that will produce gametes (or spores). These changes
will be inherited by offspring. In addition, errors in meiosis can occur because
of the incorrect segregation of chromosomes or unequal exchange of DNA during
crossing over. These changes will be passed to offspring, also, obviously, as
they are happening during the process of gamete formation. These heritable changes
are the focus of the this lecture and the next one.
The changes can occur at several
levels, changing the number of chromosomal sets, changing the number of chromosomes
in a set, changing the number and arrangement of genes on chromosomes, and changing
the nitrogenous base sequence of genes. ALL of these changes are mutations,
and we will look at each level of changes in order - from large changes in chromosome
number to small changes in nitrogenous base sequence.
For the most part, mutations occur
randomly. And as you would suspect, making random changes to a complex, functional
system like a living cell are probably going to have a negative effect. This
is particularly true for large changes, like the gain or loss of individual
chromosomes or chromosome sets. In humans and other animals with complex development
and tissue specialization, most of the large-scale changes cause such profound
problems with development that the embryo does not develop correctly and is
spontaneously aborted. Over 90% of all spontaneous abortuses (that are tested)
have chromosomal anomalies. By analogy, think about the complex system of an
automobile. If you make a large, random change to a functioning car - like taking
out the engine, or putting one big wheel on the front right axle - it is unlikely
that you will improve the functioning of the system. Small changes are probably
going to have a negative effect, also, but it is more likely that a small change
has a beneficial effect than a large change. For example, you might randomly
change the angle of the rear-view mirror every time you get in the car. Well,
sometimes you will throw it off so you can't see, but sometimes, just by chance,
you will make a small change that adjusts it to a better viewing angle - even
if it was already pretty good.
B. A Change in
Ploidy
This is the most dramatic change
possible; an addition of an entire SET of chromosomes, changing a haploid gamete
to a diploid gamete, or changing a diploid cell into a tetraploid cell.
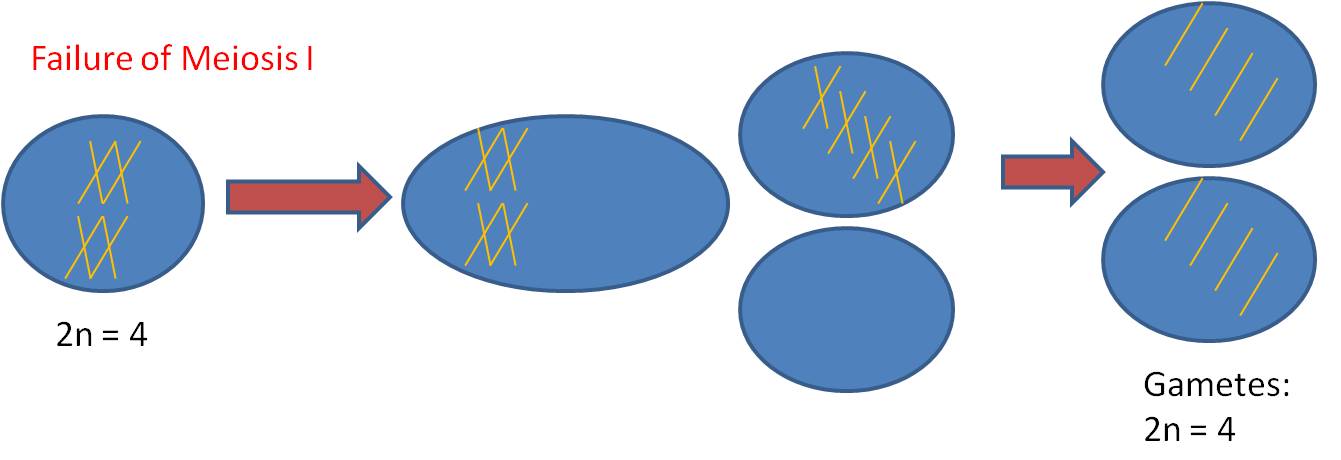 1.
Mechanism #1 - Complete failure of Meiosis
1.
Mechanism #1 - Complete failure of Meiosis
a. process
- - There is no reduction; a diploid
gamete is produced. This can occur because of a failure to divide properly
during meiosis I or meiosis II.
- - It is unlikely that this rare
event would occur in two different individuals at the same time, so typically
this diploid gamete will fertilize a normal haploid gamete, resulting in a
TRIPLOID (3n) zygote.
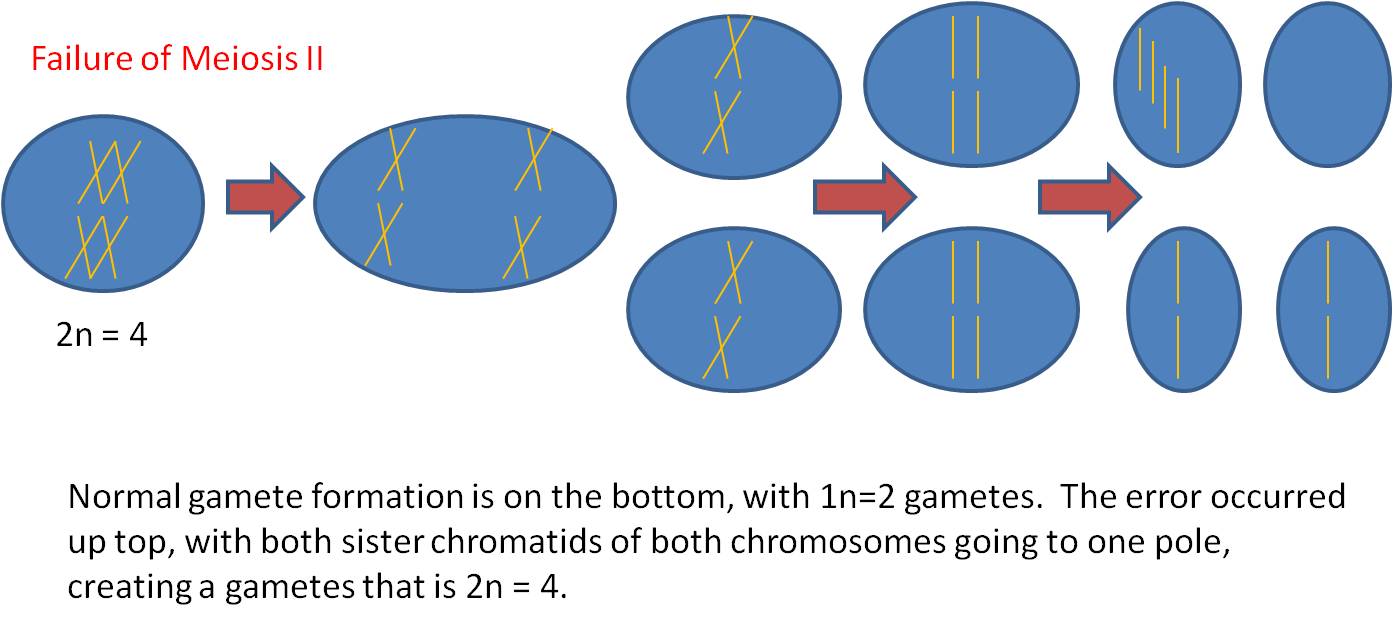
b. result
- a diploid gamete is produced.
- usually, this gamete is fertilized
by a normal haploid gamete, resulting in a triploid zygote.
- this is almost always BAD - especially
in animals - resulting in the inability of the zygote to develop correctly
and the spontaneous abortion of the embryo.
- Even if the organism can survive,
triploidy is usually a reproductive dead-end because triploids can't produce
gametes by meiosis (how do you divide 3n equally?).
- But, there are triploid species;
they have gotten around this by reproducing parthenogenetically - without
fertilization. They produce 3n gametes by mitosis,
without reduction (just like THEY were probably produced...suggesting some
possibility of a heritable aspect to this mistake). These 3n eggs develope
without fertilization into 3n adults... clones of the original female parent.
(All triploid lineages that survive in this way are females).
- So, when this happens, a new unique
lineage is created that does not exchange genes with any other group... it
is a new 'instant' species. Again,
however, this is rather rare in most higher vertebrates. Apparently, the addition
of an entire set of chromosomes disrupts normal development - probably by
changing the protein concentrations and regulatory timing in the embryo.
Study Questions:
1. Explain why females are often larger than
males, even in species in which individuals change sex.
2. Provide an example of social sex determinism.
3. What is 'gender'?
4. How do animals use sex, beside procreation?
5. Describe a circumstance where members of one
sex in animals accept the 'gender role' of the other sex.
6. Explain why sex linkage violates the priciple
of independent assortment.
7. Consider this cross: AaXBY
x AaXBXb
- How many types of gametes can the male
make with respect to these loci?
- How many phenotypes are possible in the
progeny (Assume IA and complete dominance for both loci).
- What fraction of male offspring will
express the Ab phenotype?
8. Consider these
results from the following testcross:
AaBb x aabb
F1 Phenotypes:
AB 15
Ab 22
aB 29
ab 13
Conduct a chi-square
test of independence. Using the p = 0.05 level (and critical X2
of 3.64), make a conclusion about whether the genes are linked or assort independently.
If the genes are
linked, map their positions in the heterozygous parent, showing which alleles
are on which homolog, and determining the distance between loci.
9. Describe how
polyploidy occurs.
10. Why is polyploidy
typically BAD?
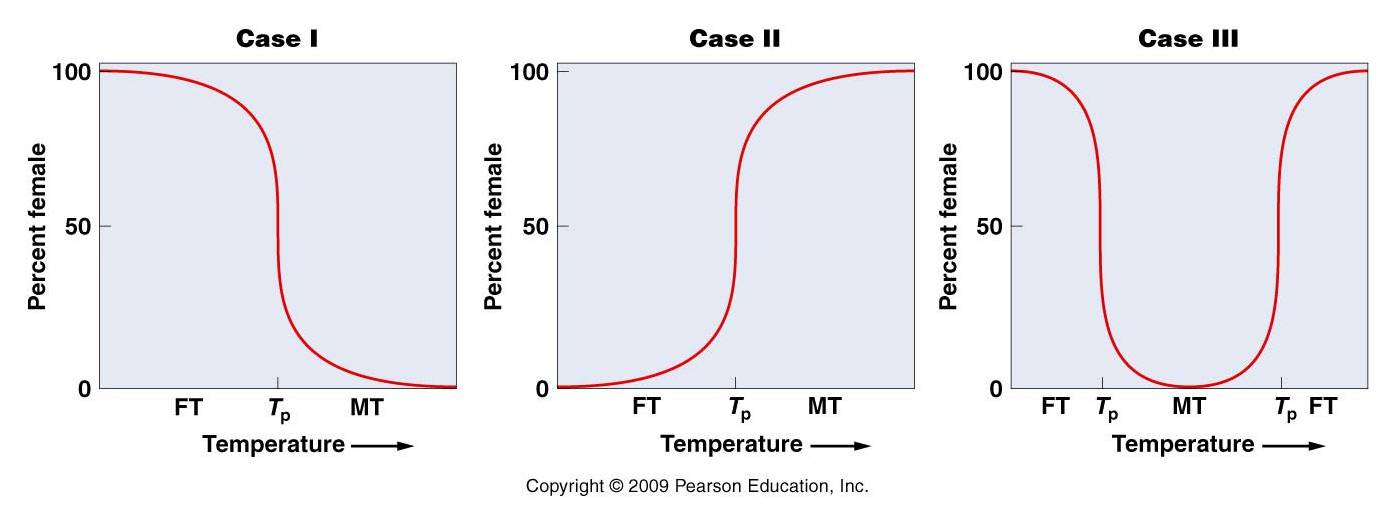 a.
temperature (in some
turtles, warm eggs develop into daughters and cool eggs into sons; in some lizards
and crocodilians, the pattern is reversed: cool = female, warm = sons). These
effects are not direct; what happens is that temperature differentially activates
sets of genes that influence certain hormonal pathways that affect the development
of gonads and genitalia. But these genes are not all on a given chromosome,
so there is no correlation between chromosomal complement or genotype and sex.
Species with temperature dependent sex determination don't change sex; sex is
determined once, by the temperature of incubation. Here
is a nice overview of the topic by Dr.
Michael Dorcas at Davidson.
a.
temperature (in some
turtles, warm eggs develop into daughters and cool eggs into sons; in some lizards
and crocodilians, the pattern is reversed: cool = female, warm = sons). These
effects are not direct; what happens is that temperature differentially activates
sets of genes that influence certain hormonal pathways that affect the development
of gonads and genitalia. But these genes are not all on a given chromosome,
so there is no correlation between chromosomal complement or genotype and sex.
Species with temperature dependent sex determination don't change sex; sex is
determined once, by the temperature of incubation. Here
is a nice overview of the topic by Dr.
Michael Dorcas at Davidson.
 probabilistic
argument.... if everyone else is male and you are the only female, then you
will mate and all of the offspring in the population will be yours (HUGE relative
reproductive success). However this reason is really unrelated to size. A second
reason is because you can adjust the QUALITY of your offspring. You can decide
to invest in all of them, or can invest in fewer and give them each more energy.
This give them a better chance of survival, which increases the likelyhood that
your genes (that are in these offspring), get passed on. So, if you are a female
plant and you have received pollen from many males, you can abort the embryos
that are doing poorly and only invest energy in embryos that are doing well.
This increases your reproductive success. And, this selective abortion doesn't
happen only in plants. European red deer (Cervus elaphus) have been
shown to selectively abort male embryos in years where resources are in short
supply. How can we understand this? Well, it is understandable in light of natural
selection and differential reproductive success in this harem-forming species.
Small male fawns grow into small male adults who won't mate - they won't acquire
a harem. Small does, however, still have a high probability of mating. So, consider
two starving, adult females that are pregnant with a male embryo and a felame
embryo, respectively. If they both bring their embryo to term and deliver it,
they will probably reduce the probability of their own survivorship (using the
last of their energy on their offspring) - so there is a cost. For the doe with
a male embryo, there is no fitness benefit; it's offspring won't mate. For the
doe with a female embryo, there will still be a reproductive payoff, as the
female offspring may still mate. Now, consider another starving doe with a male
embryo. If she aborts this embryo, she saves all the energy she would have spent
on growing that embryo to term. With thsi saved energy, she has a better chance
of surviving, herself - and reproducing, again. So, when resources are in short
supply, selection favors females that abort male embryos (and save the energy
to survive and breed again), but let female embryos develop (that are likely
to mate even though they are small). We
got a bit off track here - deer have chromosomally determined sex determination;
appreciate that this was just an example of selective abortion.
probabilistic
argument.... if everyone else is male and you are the only female, then you
will mate and all of the offspring in the population will be yours (HUGE relative
reproductive success). However this reason is really unrelated to size. A second
reason is because you can adjust the QUALITY of your offspring. You can decide
to invest in all of them, or can invest in fewer and give them each more energy.
This give them a better chance of survival, which increases the likelyhood that
your genes (that are in these offspring), get passed on. So, if you are a female
plant and you have received pollen from many males, you can abort the embryos
that are doing poorly and only invest energy in embryos that are doing well.
This increases your reproductive success. And, this selective abortion doesn't
happen only in plants. European red deer (Cervus elaphus) have been
shown to selectively abort male embryos in years where resources are in short
supply. How can we understand this? Well, it is understandable in light of natural
selection and differential reproductive success in this harem-forming species.
Small male fawns grow into small male adults who won't mate - they won't acquire
a harem. Small does, however, still have a high probability of mating. So, consider
two starving, adult females that are pregnant with a male embryo and a felame
embryo, respectively. If they both bring their embryo to term and deliver it,
they will probably reduce the probability of their own survivorship (using the
last of their energy on their offspring) - so there is a cost. For the doe with
a male embryo, there is no fitness benefit; it's offspring won't mate. For the
doe with a female embryo, there will still be a reproductive payoff, as the
female offspring may still mate. Now, consider another starving doe with a male
embryo. If she aborts this embryo, she saves all the energy she would have spent
on growing that embryo to term. With thsi saved energy, she has a better chance
of surviving, herself - and reproducing, again. So, when resources are in short
supply, selection favors females that abort male embryos (and save the energy
to survive and breed again), but let female embryos develop (that are likely
to mate even though they are small). We
got a bit off track here - deer have chromosomally determined sex determination;
appreciate that this was just an example of selective abortion.
 c.
Social Environment:
Many territorial reef fish like clown fish and wrasses have sequential hermaphrodism,
as we mentioned before. A dominant, large, mature individual (female clownfish,
male wrasse) dominates the other individuals in the school. In clownfish, all
but one of the other fish in the school remain juvenile males; the other developes
into a reproductive male who pairs with the dominant female. With the loss of
the mature dominant female, the mature male changes sex and becomes the dominant
female, one immature male developes into a mature male, and the others remain
sexually immature. In progynous wrasses, one female in a school developes into
a reproductive male who mates with the mature females. The loss of the male
stiumulates one of the females to change sex. The nature of these changes is
poorly understood, but in both cases the social dynamics in the school have
an effect. Aggressive behaviors by the dominant individual influence the hormone
concentrations in subordinate individuals, maintaining them as immatures or
in the other sex. Loss of this suppression (by the loss of the dominant individual)
releases one individual, who changes very quickly (often with associated morphological
and behavior changes) and assumes the aggressive, dominating behavior that continues
to suppress development in the other members of the school.
c.
Social Environment:
Many territorial reef fish like clown fish and wrasses have sequential hermaphrodism,
as we mentioned before. A dominant, large, mature individual (female clownfish,
male wrasse) dominates the other individuals in the school. In clownfish, all
but one of the other fish in the school remain juvenile males; the other developes
into a reproductive male who pairs with the dominant female. With the loss of
the mature dominant female, the mature male changes sex and becomes the dominant
female, one immature male developes into a mature male, and the others remain
sexually immature. In progynous wrasses, one female in a school developes into
a reproductive male who mates with the mature females. The loss of the male
stiumulates one of the females to change sex. The nature of these changes is
poorly understood, but in both cases the social dynamics in the school have
an effect. Aggressive behaviors by the dominant individual influence the hormone
concentrations in subordinate individuals, maintaining them as immatures or
in the other sex. Loss of this suppression (by the loss of the dominant individual)
releases one individual, who changes very quickly (often with associated morphological
and behavior changes) and assumes the aggressive, dominating behavior that continues
to suppress development in the other members of the school.
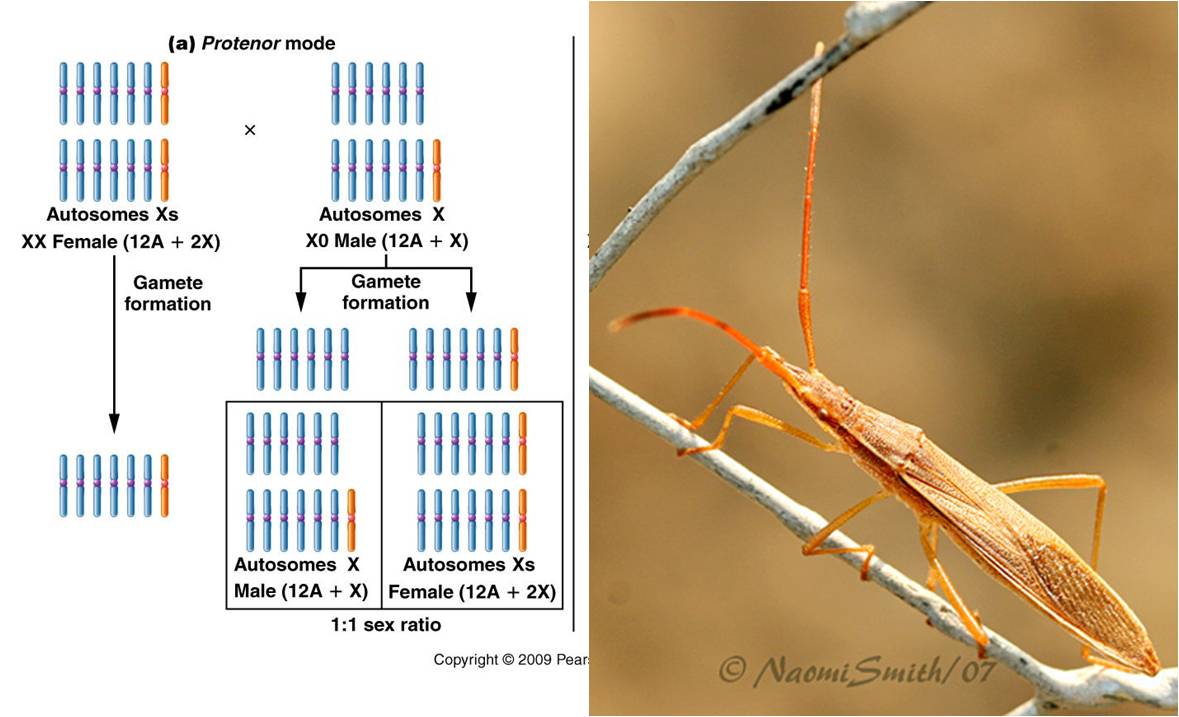 a.
Protenor (a
Homopteran... a true bug) - A single sex chromosome - 6 autosomal, no sex chromosome
(female)
a.
Protenor (a
Homopteran... a true bug) - A single sex chromosome - 6 autosomal, no sex chromosome
(female)