 b.
Islands Faunas
b.
Islands Faunas
One of the great ironies of science is that the two greatest contributions in biology - the theory of evolution and the mechanistic principles of heredity - were described independently of one another within a ten year period. It is ironic because genetics (heredity) and evolution are so critically and intimately related...heredity describes how genetic information is passed from parent to offspring. This creates relatedness patterns within families, within species among populations, and among species. These relationships are a focus of evolutionary studies. The publications marking their official, orthodox births were published only six years apart: Origin of Species (1859), and Experiments in Plant Hybridization (1865), yet it was nearly 80 years before these ideas were placed in their proper context within the Modern Synthetic Theory of evolution. In the first part of this course, we will examine these contributions in a historical context. We will also briefly describe the cellular context in which the genetic system operates.
1. Geology
2. Paleontology
3. Comparative Anatomy
4. Biogeography
a. Community
convergence
Under similar environmental conditions, we find different
species filling similar ecological niches. Outward 'form' correlates with
ecological niche (role) across entire communities. So, in Australia, marsupials
fill the role of dog-like predator, cat-like predator, burrowing animal, ant-eater,
etc. These same roles are filled by outwardly similar placental mammals in South
America. However, the similarity between a wolf (placental) and a Thylacine
(marsupial - the 'tasmanian wolf') are strictly ANALOGIES. Their underlying
structure shows them to be quite different - a wolf is more similar to a ground
hog (both placentals) in underlying structure than to a thylacine.
b.
Islands Faunas
Islands often have fewer species than a mainland - even a patch of mainland the same size. As such, the patterns and interactions are often simpler to describe and understand. For both Darwin and Alfred Russel Wallace (the other independent author of the theory of evolution by natural selection), the study of islands was critical in to the development of their ideas.
1. Distance correlates with the uniqueness of the inhabitants: the animals on the Fauklands are the same species as on the mainland, but the Galapagos fauna is composed of unique species, found nowhere else:
"The natural history of these islands is eminently curious, and well deserves
attention. Most of the organic productions are aboriginal creations, found nowhere
else; there is even a difference between the inhabitants of the different islands;
yet all show a marked relationship with those of America, though separated from
that continent by an open space of ocean, between 500 and 600 miles in width.
The archipelago is a little world within itself, or rather a satellite attached
to America, whence it has derived a few stray colonists, and has received the
general character of its indigenous productions. Considering the small size
of the islands, we feel the more astonished at the number of their aboriginal
beings, and at their confined range. Seeing every height crowned with its crater,
and the boundaries of most of the lava- streams still distinct, we are led to
believe that within a period geologically recent the unbroken ocean was here
spread out. Hence, both in space and time, we seem to be brought somewhat near
to that great fact -- that mystery of mysteries -- the first appearance of new
beings on this earth." The Voyage of the Beagle - Darwin (1839).
 2. The Galapagos fauna:
2. The Galapagos fauna:
- It was related to american fauna,
yet different: the types of animals are new world animals.... there are iguanas
like the green iguana of Central and South America, but the iguanas are different
species. So, darwin describe it as " a world within itself, or rather,
a satellite of the Americas" .... it was different, but more like the American
fauna than any other...(no chameleons, for instance, which are old world lizards...)
- It was dominated by dispersive forms. This is critical. The communities are dominated by reptiles, birds, and marine mammals. All of these organisms could MIGRATE to the islands from the mainland. (Terrestrial mammals don't migrate as well as terrestrial reptiles over open ocean. Throw a reptile in cold salty water, and: 1) its metabolism slows down (its cold), so 2) its demand for food and water decline; and 3) its scales protect it against water loss... which is why reptiles do well in the desert, too. Throw a mammal in cold salt water, and it's going to have a VERY tought time: 1) the temperature gradient between its warm body and the cold ocean is very large - in order to maintain its high body temperature against this gradient, it's metabolism has to INCREASE (to produce more heat to compensate for the heat lost to the environment). This increased metabolic demand will INCREASE the need for food and water... that's probably in pretty short supply in the open ocean; and 2) water is lost quickly from the skin to the salty ocean once the fur is wet... so, mammals are more likely to starve or die of exposure than reptiles.
- So, the islands are dominated by dispersive forms, and this suggests they came from America. But if they came from America, WHY ARE THEY DIFFERENT SPECIES THAN THOSE IN AMERICA? They must have changed since their arrival.
- There are even differences between
species on different islands. On the 14 species of finches - "Seeing this gradation
and diversity of structure in one small, intimately related group of birds,
one might really fancy that from an original paucity of birds in this archipelago,
one species had been taken and modified for different ends." The Voyage
of the Beagle - Darwin (1839)  VIDEO
VIDEO
- Galapagos Mockingbirds Darwin thought he had one, very variable species. He
sent specimens to John Gould, the premiere ornithologist in England at the time.
On Darwin's return to England, he found out that Gould recognized the mockingbird
specimens as belonging to 4 separate species. This made Darwin consider that
maybe the variation WITHIN a species could be continuous with the variation
BETWEEN species.... maybe varieties within a species could gradually become
so different from one another that they would eventually become different species.
VIDEO
5. Argument For Evolution as a Historical Fact:
Premise 1: Species that are alive today are different from those that have lived previously.
Premise 2: Spontaneous Generation is refuted, so organisms only come from other organisms.
Conclusion 1: Thus, the organisms alive today must have come from those pre-existing, yet different, species.
Conclusion 2: There must have been change through time (evolution).
Conclusion 3: The fossil record, vestigial organs, and homologies are all suggestive of descent from common ancestors.Below, the figure from The Origin of Species that shows Darwin's idea of descent from common ancestors.
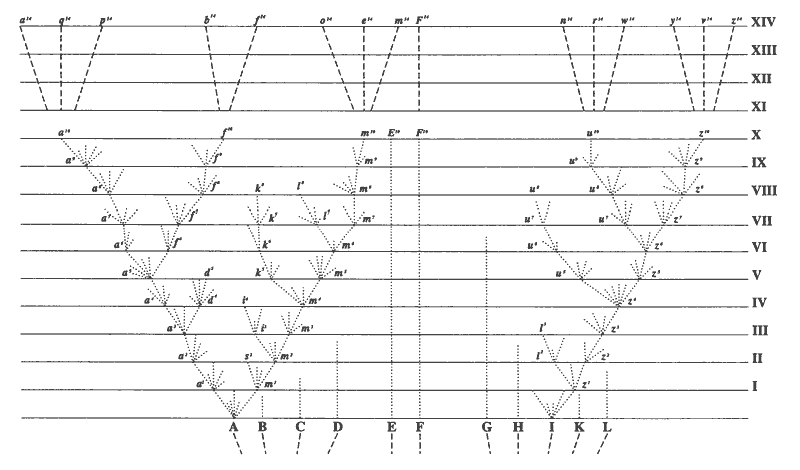
So, if species do change over time (evolve), the next question is "How?" How does this change occur?
1. Transitional Observations
 a.
Domesticated animals:
a.
Domesticated animals:
As a result of selective breeding, humans have taken certain species and modified them tremendously. So, from an ancestral population of wolves, we have created chihuahuas and St. Bernards. Now, there were never any Chihuahua sized wolves running around - we created this variability by progressively breeding smaller dogs with one another. Now, we have two groups, Chihuahuas and St. Bernards, that can't easily be bred together. So, we have created separate biological groups, that could be called different species. Darwin was aware of many 'breeds' of pigeons, too. These were called varieties, almost assuredly descended from a common ancestral population of rock doves (left). The KEY to the production of new varieties by humans is that humans only allowed certain organisms to breed. So, only particular traits were passed to the next generation. Darwin wondered if there was a mechanism that could do the SAME thing in nature (only allow certain organisms in a population to breed), and thereby explain the natural changes seen in the fossil record (radiational patterns of multiple species in one strata necessarily coming from fewer species in earlier strata). and implied by island faunas. Just because man does it purposefully, by design and with aforethought, doesn't mean that nature can't do it WITHOUT purpose. We put water in the freezer to PURPOSEFULLY make ice cubes... but water freezes naturally, as well - without a purpose, but with the same NATURAL CAUSE (loss of heat).
 b. Reading Malthus:
b. Reading Malthus:
In 1838, Darwin read a book published in 1798 by Thomas Malthus called Essay on the Principle of Population. Malthus was a british aristocrat who was concerned that the british aristocracy might be overthrown like the French had been 10 years before (1788). He realized that all populations have the capacity to grow exponentially, but that resources are finite. As such, there will eventually be a "struggle for existence" (Darwin's words). Malthus was worried that the british population would continue to grow, and that poverty and famine would lead to revolution.
Darwin read Malthus, and wrote in his autobiography:
“In October 1838,
that is, fifteen months after I had begun my systematic enquiry, I happened
to read for amusement Malthus on Population and being well prepared to appreciate
the struggle for existence which everywhere goes on from long-continued observation
of the habits of animals and plants, it at once struck me that under these circumstances
favourable variations would tend to be preserved, and unfavourable ones to be
destroyed. The result of this would be the formation of new species. Here, then,
I had at last got a theory by which to work; but I was so anxious to avoid prejudice,
that I determined not for some time to write even the briefest sketch of it.
In June 1842 I first allowed myself the satisfaction of writing a very brief
abstract of my theory in pencil in 35 pages; and this was enlarged during the
summer of 1844 into one of 230 pages, which I had fairly copied out and still
possess.” - The Autobiography of Charles Darwin
1809-1882 (Barlow 1958).
 -
If there are limited resources of food, shelter, and mates, and if organisms
in a population vary, then as a consequence of this variability, some will be
more likely to gain the resources than others, and will thus be more likely
to mate. So, the environment will 'select' which animals in a population will
mate (the ones best able to acquire the limiting resources)... and these traits
that work to gain resources in this environment will be passed on at high frequency
to the next generation.
-
If there are limited resources of food, shelter, and mates, and if organisms
in a population vary, then as a consequence of this variability, some will be
more likely to gain the resources than others, and will thus be more likely
to mate. So, the environment will 'select' which animals in a population will
mate (the ones best able to acquire the limiting resources)... and these traits
that work to gain resources in this environment will be passed on at high frequency
to the next generation.
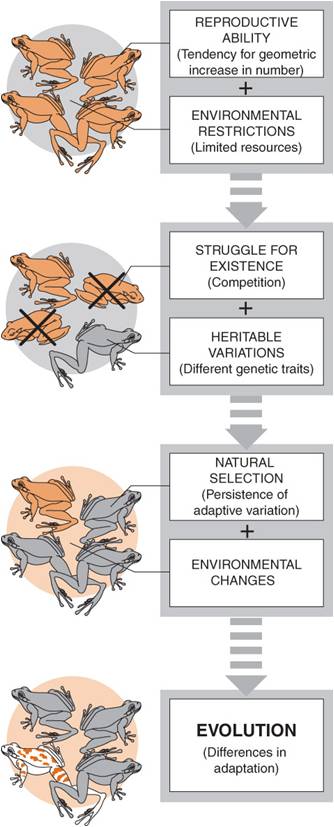
2. Natural Selection: (Know this. Understand it. You WILL be asked to outline NS in this very form.)
P1: Populations over-reproduce (Malthus)
P2: resources are finite (Malthus)
C1: Eventually, a population will grow until it becomes limited by its resources. At that time, their will be a "struggle for existence" and most offspring produced will die. (Malthus)
P3: Individuals in a population vary, and some of this variation is heritable (Darwin - observations and animal/plant breeding)
C2: Variations will not have the same probability of survival and reproduction in a particular environment; those well-suited to the environment will be more likely to survive and reproduce than others, passing on the genes for these adapted traits. There will be "Differential Reproductive Success" (Observations, breeding).
C3: Over time, adaptive traits will accumulate and the characteristics in a population will change. This is lineage evolution. (Like change in horse toes in a sequence of fossil species, or like the change in the chihuahua lineage from the ancestral wolves).
Corollary:
Two sub-populations, separated in different environments, would be selected
for different traits and may subsequently lose the capacity to interbreed. At
this point, they are different biological species. This is Speciation and Radiational
Evolution. (like the production of different Finches, mockingbirds, etc. on
different islands in the galapagos, and like the radiation of St. Bernards AND
chihuahua's, which diverged from one another over time).
Darwin here provides a natural explanation for why purposeful structures and behaviors occurs in nature. Through some process unknown to him, variation arises in natural populations. These varieties differ in terms of functional efficiency in a common environment; so some improving an organisms probability of surviving and mating than others. Organisms with these beneficial traits will leave more offspring, and the frequencies of these beneficial characteristics will increase through time - much as humans select for smaller and smaller dogs. He ends The Origin of Species (1859) like this:
 "It
is interesting to contemplate an entangled bank, clothed with many plants of
many kinds, with birds singing on the bushes, with various insects flitting
about, and with worms crawling through the damp earth, and to reflect that these
elaborately constructed forms, so different from each other, and dependent on
each other in so complex a manner, have all been produced by laws acting around
us. These laws, taken in the largest sense, being Growth with Reproduction;
Inheritance which is almost implied by reproduction; Variability from the indirect
and direct action of the external conditions of life, and from use and disuse;
a Ratio of Increase so high as to lead to a Struggle for Life, and as a consequence
to Natural Selection, entailing Divergence of Character and the Extinction of
less-improved forms. Thus, from the war of nature, from famine and death, the
most exalted object which we are capable of conceiving, namely, the production
of the higher animals, directly follows. There is grandeur in this view of life,
with its several powers, having been originally breathed into a few forms or
into one; and that, whilst this planet has gone cycling on according to the
fixed law of gravity, from so simple a beginning endless forms most beautiful
and most wonderful have been, and are being, evolved". -
The Origin of Species (Darwin 1859).
"It
is interesting to contemplate an entangled bank, clothed with many plants of
many kinds, with birds singing on the bushes, with various insects flitting
about, and with worms crawling through the damp earth, and to reflect that these
elaborately constructed forms, so different from each other, and dependent on
each other in so complex a manner, have all been produced by laws acting around
us. These laws, taken in the largest sense, being Growth with Reproduction;
Inheritance which is almost implied by reproduction; Variability from the indirect
and direct action of the external conditions of life, and from use and disuse;
a Ratio of Increase so high as to lead to a Struggle for Life, and as a consequence
to Natural Selection, entailing Divergence of Character and the Extinction of
less-improved forms. Thus, from the war of nature, from famine and death, the
most exalted object which we are capable of conceiving, namely, the production
of the higher animals, directly follows. There is grandeur in this view of life,
with its several powers, having been originally breathed into a few forms or
into one; and that, whilst this planet has gone cycling on according to the
fixed law of gravity, from so simple a beginning endless forms most beautiful
and most wonderful have been, and are being, evolved". -
The Origin of Species (Darwin 1859).
Chapter Six in "The Origin of Species" is entitled: Difficulties
on Theory
Darwin starts the chapter by writing:
"Long before having arrived at this part of my work, a crowd of difficulties will have occurred to the reader. Some of them are so grave that to this day I can never reflect on them without being staggered; but, to the best of my judgment, the greater number are only apparent, and those that are real are not, I think, fatal to my theory."
These dilemmas are:
1. How can we explain the existence of complex characteristics composed of mutually dependent parts?
- Paley's "watchmaker analogy" (1798) and the eye - the argument of design. Some elements of nature seem so complex, and composed of mutually dependent parts, that it does not seem possible that they could arise by sequential innovations. For instance, for Paley, half an eye won't work. For modern Intelligent Design proponents, the cell is called "irreducibly complex" - suggesting that it can't come from any simpler precursor - everything has to be there. (Of course, for philosophical reasons, this is about as unscientific as it gets. The idea that something is simply "too complex" to understand thwarts curiosity and stifles TRUE scientific inquiry, which is fundamentally reductionistic - as we have discussed.)
- Darwin's Solution:
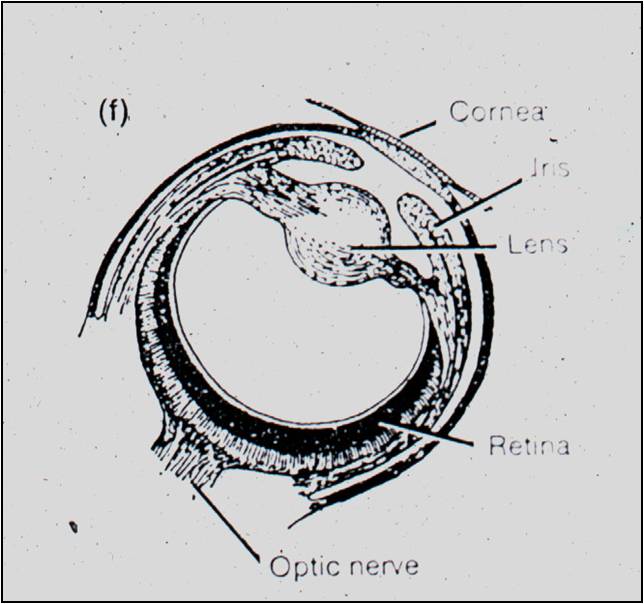 "To
suppose that the eye, with all its inimitable contrivances for adjusting the
focus to different distances, for admitting different amounts of light, and
for the correction of spherical and chromatic aberration, could have been formed
by natural selection, seems, I freely confess, absurd in the highest possible
degree. Yet reason tells me, that if numerous gradations from a perfect and
complex eye to one very imperfect and simple, each grade being useful to its
possessor, can be shown to exist; if further, the eye does vary ever so slightly,
and the variations be inherited, which is certainly the case; and if any variation
or modification in the organ be ever useful to an animal under changing conditions
of life, then the difficulty of believing that a perfect and complex eye could
be formed by natural selection, though insuperable by our imagination, can hardly
be considered real." - The Origin of Species (Darwin 1859).
"To
suppose that the eye, with all its inimitable contrivances for adjusting the
focus to different distances, for admitting different amounts of light, and
for the correction of spherical and chromatic aberration, could have been formed
by natural selection, seems, I freely confess, absurd in the highest possible
degree. Yet reason tells me, that if numerous gradations from a perfect and
complex eye to one very imperfect and simple, each grade being useful to its
possessor, can be shown to exist; if further, the eye does vary ever so slightly,
and the variations be inherited, which is certainly the case; and if any variation
or modification in the organ be ever useful to an animal under changing conditions
of life, then the difficulty of believing that a perfect and complex eye could
be formed by natural selection, though insuperable by our imagination, can hardly
be considered real." - The Origin of Species (Darwin 1859).
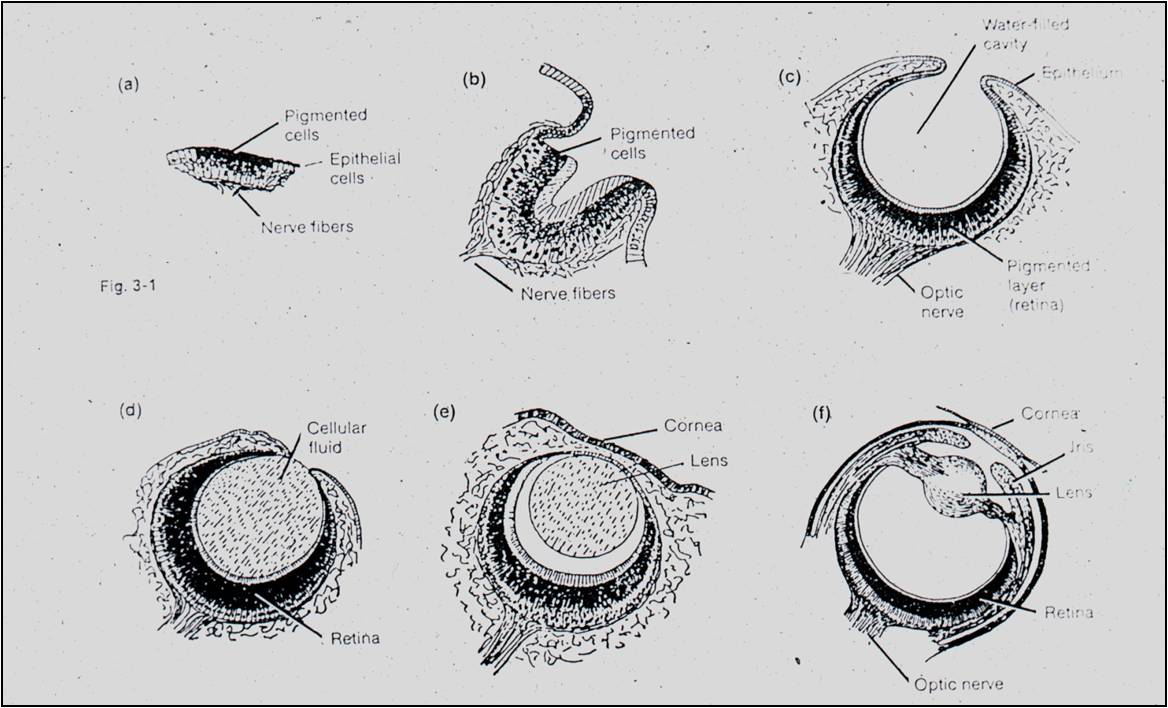 Well,
Darwin then describes the photoreceptive organs in molluscs. They range from
a sheet of receptive cells (a naked retina) to a cupped retina, to a lensless
pinhole eye, to an eye with a lens, to a camera eye. If this sequence exists
today, it is possible that this sequence could have existed through time, as
a sequential evolutionary series in eye evolution. Half an eye CAN work, if
the half you evolve first is the retina. The following videos from David
Attenborough and Richard
Dawkins are particularly good at describing the likely evolutionary scenario.
Well,
Darwin then describes the photoreceptive organs in molluscs. They range from
a sheet of receptive cells (a naked retina) to a cupped retina, to a lensless
pinhole eye, to an eye with a lens, to a camera eye. If this sequence exists
today, it is possible that this sequence could have existed through time, as
a sequential evolutionary series in eye evolution. Half an eye CAN work, if
the half you evolve first is the retina. The following videos from David
Attenborough and Richard
Dawkins are particularly good at describing the likely evolutionary scenario.
2. Where are all the intermediates?
“…why, if species have descended from other species by insensibly fine gradations, do we not everywhere see innumerable transitional forms? Why is not all nature in confusion instead of the species being, as we see them, well defined? … as by this theory innumerable transitional forms must have existed, why do we not find them embedded in countless numbers in the crust of the earth?” The Origin of Species (Darwin 1859).
Darwin was well aware that his uniformitarian hypothesis would require populations to change continuously and gradually through time. And although Lamarck's work on molluscs showed this type of change, most other lineages were best described as discontinuous and incomplete. And of course, and most importantly, the hypothesis of common ancestry predicts the existence of transitional species, representing the first evolutionary steps in the evolution of a major new group. No transitional fossils had been discovered by 1859.
There was another set of 'intermediates' that Darwin's hypothesis predicted - internediates between existing species. In other words, what would prevent the intermediates in a temporal sequence to leave descendants of their intermediate forms, thus making a continuous sequence of LIVING species between sister taxa? This continuity would make resolving one species from another almost impossible ("nature in confusion"). Darwin addressed this issue first. He said that selection, itself, would solve this issue. A more efficient descendant, with new adaptive traits, would competitively eliminate less adapted, ancestral forms:
“As natural selection acts solely by the preservation of profitable modifications, each new form will tend in a fully-stocked country to take the place of, and finally to exterminate, its own less improved parent or other less-favoured forms with which it comes into competition. Thus extinction and natural selection will, as we have seen, go hand in hand. Hence, if we look at each species as descended from some other unknown form, both the parent and all the transitional varieties will generally have been exterminated by the very process of formation and perfection of the new form.” – The Origin of Species (Darwin 1859)
 With
respect to intermediate and transitional fossils, Darwin suggested that the
fossil record was incomplete. Fossilization was a very rare event, and thus
it would be very unlikely for a representative of each species in a lineage
to have a representative preserved. However, he hope that, in time, a more complete
picture (and some truly transitional fossils) would be discovered. In 1861,
he was pleased by the discovery of Archeopteryx lithographica in Bavaria.
When previous fossils of this species had been unearthed, they were classified
as reptiles because they had teeth, fingers, and a long bony tail. But, when
this fossil (and 9 others since) were pulled from the very fine sedimentary
deposits of Bavaria, the impression of feathers were seen. So, here is an organism
with a very odd combination of traits - a lizard skeleton covered by feathers.
With
respect to intermediate and transitional fossils, Darwin suggested that the
fossil record was incomplete. Fossilization was a very rare event, and thus
it would be very unlikely for a representative of each species in a lineage
to have a representative preserved. However, he hope that, in time, a more complete
picture (and some truly transitional fossils) would be discovered. In 1861,
he was pleased by the discovery of Archeopteryx lithographica in Bavaria.
When previous fossils of this species had been unearthed, they were classified
as reptiles because they had teeth, fingers, and a long bony tail. But, when
this fossil (and 9 others since) were pulled from the very fine sedimentary
deposits of Bavaria, the impression of feathers were seen. So, here is an organism
with a very odd combination of traits - a lizard skeleton covered by feathers.
- IMPORTANT: Now, the existence of this odd animal is not the only thing that bears on evolution. Obviously, evolution does predict the existence of intermediates. But even more so, it also predicts WHEN in the fossil record this species should have lived. For instance, If Archeopteryx is a transitional species, it has to be after reptiles appear in the fossil record, and before true birds. It can't just be anywhere and be a transitional species between these groups. Well, it is just where evolutionary theory predicts it should be; after other reptiles and before all true birds. We will see lots of intermediates later in the course, when we look at post-Darwinian developments.
3. How is heritable variation produced?
Darwin's genius was seeing the importance of heritable variation; it would lead to differential reproductive success and evolutionary change. However, he did not understand how this variation was produced (mutation and recombination) or inherited (meiosis and heredity). He knew how important this was for his theory, and he was a real scientific student of hybridization experiments. One of the most difficult chapters in The Origin of Species is his extensive summary of patterns of hybridization across the animal and plant kingdoms. It is obvious that he hoped to find patterns in comparing these disparate studies that would reveal a mechanism of heredity. Unfortunately, they did not. As such, Darwin relied on those old Lamarckian ideas of "inheritance of acquired traits" (described as the effects of the 'external conditions of life') and "use and disuse":
"These laws, taken
in the largest sense, being Growth with Reproduction; Inheritance which is almost
implied by reproduction; Variability from the indirect
and direct action of the external conditions of life, and from use and disuse;
a Ratio of Increase so high as to lead to a Struggle for Life, and as a consequence
to Natural Selection…". - The Origin of Species
(Darwin 1859).
Darwin proposed the existence
of particulate 'gemmules' that were present in every cell of the body. Changes
in the body changed these gemmules somehow, and before mating these gemmules
migrated to the reproductive tissue for transmission to the next generation
- explaining how acquired traits could be inherited. He even did experiment
to test his idea, which he falsified. Yet, he had no other ideas. He
died not appreciating the insights that had been made by Mendel in Austria in
1865.
E. A Summary of Darwinian Evolution:
Darwin's view of evolution can be summarized as follows:
Source of variation: UNKNOWN
but variation is observable
Causes of Evolutionary Change:
(factors that cause change in natural populations): Natural Selection
In this unit, we will examine how DNA is transferred through time, we will describe the process of sexual reproduction, and we will solve Darwin's Dilemma regarding the source of heritable variation. In addition, we will examine how the activity of genes is regulated - a process that results in the coordinated specialization of cell types and tissues through time (development).
 A.
Mendel's Life (from Villanova
University)
A.
Mendel's Life (from Villanova
University) Johann Mendel was born in Moravia (now the Czech Republic) on July 22, 1822, into a poor farming family. He received an excellent education at the local schools, and entered the Augustinian order at the St. Thomas monastery in Brunn (now Brno) in1843. He was ordained in 1847 and was given the name 'Gregor'. The monastery sent him the the University of Vienna for two years (1851-1853), where he trained to be a teacher in physics and mathematics. After returning to Brunn, he began his experiments and taught part-time. He presented his work at scientific meetings in 1865, and the work was published in the proceedings of the meetings in 1866. His paper, Versuche über Pflanzen-Hybriden ("Experiments in Plant Hybridization,") provided one othe first quantitative examinations of a biological phenomena. The conclusions and implications of the work went largely unnoticed for 30 years, until the work was cited by three botanists independently working on plant hybridization in 1900. Mendel became the Abbot of the monastery, and his administrative duties put an end to his career as an experimental biologist. Mendel died in Brünn on 6 January 1884. Before his death, he stated, "My scientific labors have brought me a great deal of satisfaction, and I am convinced that before long the entire world will praise the result of these labors." His conviction came to pass, as he is now recognized as the founding father of genetics.
It was common knowledge that offspring looked alot like their parents. This observation was especially obvious and important in the agrarian societies of the pre-industrial age, where people saw that certain traits in domesticated plants, domesticated animals, and humans 'ran in families'. In fact, this was not just an idle issue. Understanding how heredity worked was an important economic question to these farmers, who were trying to breed more valuable and productive crops and livestock.
1. Preformationist Ideas
 One
school of thought - the "preformationist" school, thought that a miniature
human lived within either the egg (the 'ovist' school), or the sperm ('the homunculan'
school). This offspring laid in a state of suspended animation until the sperm
stimulated the egg to grow (for the ovist school), or the sperm was placed in
the fertile womb to grow ('homunculan' school). Of course, both of these ideas
had serious flaws. If the offspring was really just a product of one sex, then
why did the offspring have characteristics of both parents? Some theologians
saw interesting implications, too. If all generations were preformed, then each
generation must be nested within the egg or sperm of the preceeding generation
- also already preformed. The generations of humans would be like a set of russian
dolls, one nested with the other. At some point, it would seem that nature would
reach the limit of how small a preformed person could be - and that would be
the last generation (which would be followed by the apocalypse).
One
school of thought - the "preformationist" school, thought that a miniature
human lived within either the egg (the 'ovist' school), or the sperm ('the homunculan'
school). This offspring laid in a state of suspended animation until the sperm
stimulated the egg to grow (for the ovist school), or the sperm was placed in
the fertile womb to grow ('homunculan' school). Of course, both of these ideas
had serious flaws. If the offspring was really just a product of one sex, then
why did the offspring have characteristics of both parents? Some theologians
saw interesting implications, too. If all generations were preformed, then each
generation must be nested within the egg or sperm of the preceeding generation
- also already preformed. The generations of humans would be like a set of russian
dolls, one nested with the other. At some point, it would seem that nature would
reach the limit of how small a preformed person could be - and that would be
the last generation (which would be followed by the apocalypse).
2. Epigenetic Ideas
The other major school of thought promoted the "epigenetic" idea. Here, the egg does not actually contain a small individual, but rather only the potential to grow and develop into an individual. This idea had much less support, because there were no ideas about how this could happen; the growth of a small human was alot easier to understand than the development of organs and organ systems from 'nothing' - or at least, from things that weren't organs or organ systems! However, the epigenetic idea had one major benefit - it could explain why offspring often expressed characteristics of both parents. If both parents contributed 'stuff' to this epigenetic mass of potential, then the developing offspring could inherit traits from both parents. Because semen is a fluid, it seemed likely that the genetic information was a fluid. The 'mixing' of the heredity fluids from two parents might explain, sort of, why the offspring expressed a mixture of traits from both parents.
There was an additional observation that was very problematic with this blending idea, however. Most people were well aware that some traits 'skipped a generation', or even many generations. The reappearance of an ancestral trait (" you have your great-grandmother's nose") was know as a 'sport'. If the hereditary information from two parents were fluids that mixed, like black and white paint, it seemed difficult to explain how, after mixing black and white into grey, you could get some pure white paint back out of the mixture in a later generation.
 Mendel
set out to test these alternate hypotheses by breeding organisms from different
'pure-breeding' stocks together. In other words, Mendel conducted hybridization
experiments, forming hybrids by crossing strains that bred true for particular
characteristics. He selected the common garden pea, Pisum sativum,
as his study organism. He chose this plant because there were alot of pure-breeding
varieties already available; farmers had selected for different characteristics,
creating strains that produced round peas, wrinkled peas, yellow peas, green
peas, tall plants, short plants, etc. Indeed, Mendel reports that he received
34 different strains from farmers. In addition, because peas were fast growing
and fecund (produced lots of offspring - each pea is a separate offspring),
he could generate a lot of data in a matter of months. (Some characters, however,
like flower color or plant height, could only be determined in the following
year, when the offspring grew up and exhibited the trait.) Being trained in
mathematics and probability, he knew that a large sample would provide a 'truer'
indication of a pattern than a small sample.
Modern geneticists have chosen other study organisms - like fruit flies, mice,
and bacteria - for the same reason.
Mendel
set out to test these alternate hypotheses by breeding organisms from different
'pure-breeding' stocks together. In other words, Mendel conducted hybridization
experiments, forming hybrids by crossing strains that bred true for particular
characteristics. He selected the common garden pea, Pisum sativum,
as his study organism. He chose this plant because there were alot of pure-breeding
varieties already available; farmers had selected for different characteristics,
creating strains that produced round peas, wrinkled peas, yellow peas, green
peas, tall plants, short plants, etc. Indeed, Mendel reports that he received
34 different strains from farmers. In addition, because peas were fast growing
and fecund (produced lots of offspring - each pea is a separate offspring),
he could generate a lot of data in a matter of months. (Some characters, however,
like flower color or plant height, could only be determined in the following
year, when the offspring grew up and exhibited the trait.) Being trained in
mathematics and probability, he knew that a large sample would provide a 'truer'
indication of a pattern than a small sample.
Modern geneticists have chosen other study organisms - like fruit flies, mice,
and bacteria - for the same reason.
1. Monohybrid Experiments (single traits)
Mendel reported the results of seven parallel experiments. In each experiment, he cross plants that had different expressions of a particular trait. So, in the third set of trials, he bred plants that 'bred true' for white flowers (when bred with themselves) with plants that produced purple flowers. For this experiment, he did 35 cross-fertilizations with ten plants.
a. Reciprocal parental crosses tested the ovist and homunculan schools:
A very important methodological aspect of the experiment was that he conducted reciprocal crosses between stocks. So, he placed pollen pure-breeding white flowers on the stigma of purple-flowers, and pollen from purple flowers on the stigmas of white flowers. So, by comparing the results of these reciprocal crosses, he could see if the inheritance of the trait (flower color) was associated with the sex of the parent. If the homunculan school was correct, then the offspring should bear the trait expressed in the stock that donated the pollen. If the ovist school was correct, then the offspring should bear flowers the color of the strain that received the pollen and donated the egg (in the ovule).
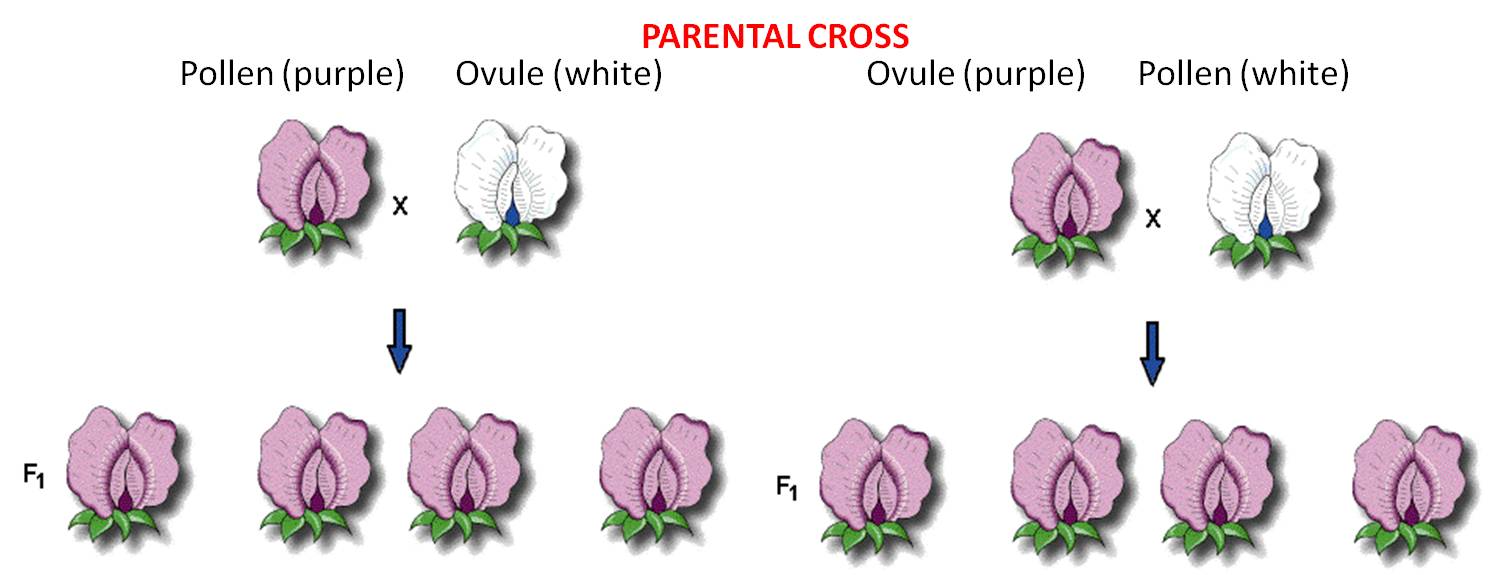 These 'pure-breeding'
stocks were called the 'parental generation'.
These 'pure-breeding'
stocks were called the 'parental generation'.
pollen (male) from Purple flowers X ovule (female) from White flowers
AND
pollen (male) from White flowers X ovule (female) of Purple flowers
These crosses produced offspring (peas). The first generation of 'hybrids' produced from pure-breeding parental strains is called the F1 (for 'first 'filial') generation. He planted and grew the peas produced from both crosses and observed their flower color. In both reciprocal crosses, all the plants produced purple flowers. These results falsified both the homunculan and ovist schools; both hypotheses predicted that plants from one or the other cross would yield white flowers. Indeed, Mendel realized that these results suggested that both parents had to contribute hereditary factors for this trait; the purple offspring from the first reciprocal demonstrated that the male contributes, and the purple offspring in the other reciprocal demonstrated that the female contributes, too. But if both parents contributed hereditary information, what happened to the white information contributed by the other parent in each cross? Mendel hypothesized that the contribution was present in the offspring, but not expressed. He coined the term 'dominant' for the trait expressed in the F1, and 'recessive' for the trait he thought was there, but hidden. He became more confident in these hypotheses when all seven traits that he studied showed the same pattern.
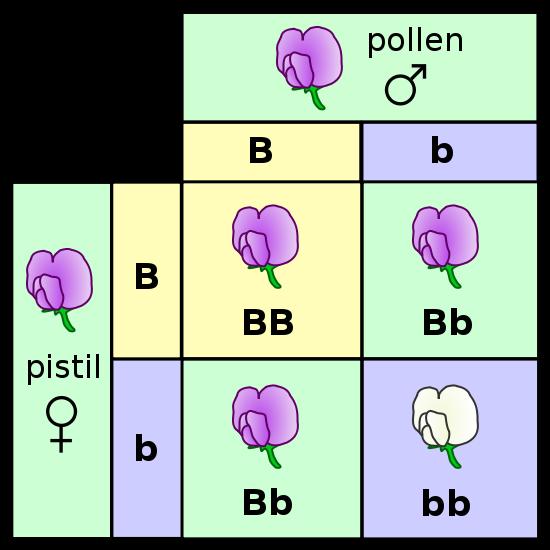 b.
Crossing the F1 hybrids tested the blending hypothesis:
b.
Crossing the F1 hybrids tested the blending hypothesis:
He allowed the F1 plants to fertilize themselves, essentially performing an F1 x F1 cross. He makes reference to the fact that, in many species and for many traits, the hybrids (F1's) exhibit a triat intermediate to the two parental types. This intermediate result might be expected of a blending pattern (white paint and black paint making grey), but one trait dominating the other could also be produced from a blending mechanism, too. One thing you would not expect from a blending of fluids, however, would be the re-expression of the recessive 'white' trait. Grey paint mixed with grey paint should not produce white paint.
The peas from these crosses were grown and the flowers were observed. Of the 929 plants that he grew from these seeds, 705 bore violet flowers and 224 had white flowers; a ratio of 3.15:1. In fact, all seven traits produced ratios near a 3:1 ratio in the F2 generation (offspring of F1 x F1 crosses). He recognized the importance of the 3:1 ratio when combining two things. It reminded him of a simple binomial expansion (a + b)2 = 1aa + 2ab + 1bb. If an organism has 2 different types of hereditary units (a = purple and b = white) for a given trait and it mates with itself, then there are three combinations that are possible in the offspring (aa, ab, bb). And, they should occur in a 1/4:2/4:1/4 ratio. If, as a consequence of dominance, the progeny with different particles (ab) only express one - the 'a' (purple) - then they will look like the aa's and 'a' (purple) offspring will occur 3/4 of the time and 'b' (white) offspring will occur only 1/4 of the time.
c. Mendel proposed four postulates (hypotheses) to explain these data:
- genetic info is unitary or 'particulate' - it is not a fluid that blends
- each oganism has two unit factors (we call these 'genes' now) for each trait
- if an organism has different factors for a given trait, one is expressed (dominant) over the other (recessive)
- the two genes for a given trait separate and go into separate gametes during gamete formation. Subsequent fertilization (fusion of gametes) is random. This is called Mendels First Principle: Principle of Segregation
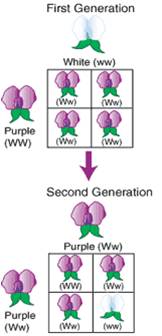 d.
How his hypothesis explains his observations:
d.
How his hypothesis explains his observations:
Parentals: WW (Purple) x ww (white) .... (and the reciprocal)
Gametes: W w
F1 offspring:
Genotypic Ratio: 100% Ww (heterozygous)
Phenotypic Ratio: 100% Purple
F1 x F1 Cross: Ww x Ww
Gametes:
W and w W
and w
F2 offspring:
Genotypic Ratio: 1/4 WW, 2/4 Ww, 1/4 ww Some Terms:
Phenotypic Ratio: 3/4 Purple, 1/4 White
Genotype = the type of genes an organism has, like Ww.
Genotypic Ratio = is the fractional representation of different
genotypes in a group, such as 1/4WW : 1/2Ww : 1/4 ww
Phenotype = the characteristic or trait that is expressed,
such as 'Purple'. Usually, this is represented by the gene that causes the trait,
such as 'W' in the example, above.
Phenotypic Ratio = the fractional representation of different
phentypes in a group, such as 3/4W : 1/4 w.
2. Monohybrid Test Crosses
If the hypotheses are correct, Mendel reasoned, then the F1 has a gene for the recessive 'white' trait that is not expressed. How could he determine whether it was there or not? He realized that he could answer this question if he crossed it to the recessive white parent: (ww). The logic is this: the homozygous recessive parent can only pass recessive alleles to the offspring. Whatever the offspring receives from the OTHER parent will determine the expression of the trait. It the offspring receives a dominant allele for purple from the other parent, then the offspring (Ww) will express purple flowers. But, if the other parent is hiding a recessive allele and passes that to the offspring, then the offspring will have the homozygous recessive genotype (ww) and will produce white flowers.
Mendel predicted that, under this hypothesis, the phenotypic ratio in the offspring would be: 50% purple (W): 50% white (w). The results supported his hypothesis, and supported his postulates. Replication of this results across all seven traits gave strong confirmation that he had described a general pattern of heredity.
More
Terms:
Homozygous: A zygote is a fertilized egg that has received
genes from both parents. If the genes received from both parents for a given
trait are the same, then this zygote (and the organism that develops from it)
is homozygous for this trait. If the zygote has received dominant genes
for the trait from both parents, it is homozygous dominant (WW); if
it has received the recessive allele from both parents, it is homozygous
recessive (ww).
Heterozygous: If the zygote receives different alleles from
the parents for a given trait, then the zygote is heterozygous for
that trait (Ww).
Alleles are different 'forms' of a gene, that affect a given
trait - like flower color - in different ways ('W'and 'w' are alleles for flower
color).
3. Dihybrid Experiments
Most animal and plant breeders had noticed that some traits seem to be inherited together. Indeed, in his own experiments, Mendel saw that flower color was always correlated with the color of the seed coat, such that seeds with white seed coats grew into plants with white flowers, and seed with brown seed coats grew into plants with purple flowers. Having demonstrated that his seven traits all exhibited the same pattern of inheritance, he now set out to determine whether these traits were inherited in a dependent or independent manner. This is a very insightful question. For example, he had already demonstrated that yellow pea color was dominant to green, that round pea shape was dominant to wrinkled, and that F1xF1 crosses for each trait produced a 3:1 ratio in the offspring. Well, maybe these traits are inherited in the same way because they are governed by the SAME heritable unit factor (gene)... and that's why they produce the same pattern. If yellow and round were caused by the same gene, then of course they would be inherited in the same way. Or, maybe they are governed by different unit factors that have a pattern of heredity that is related to one another... maybe they are governed by different factors that "travel" from parent to offspring together - yellow always traveling with round, for instance. This had important practical applications, too. A sheep farmer would profit from sheep that produce both good wool and good milk (for feta or roquefort cheese) from his sheep herd. But, maybe the trait of quality wool is inherited with the trait of poor milk production. In that case, they might not want to strongly select for great wool quality, as milk production would suffer. It might be best to maintain a variable population that produces good wool and adequate milk. Every organism expresses a combination of traits, so addressing how combinations of genes are inherited is a very important question.
a. Parental crosses of stocks breeding true for two traits
First, Mendel bred plants together and created stocks that bred true for two traits: seed color and seed shape. He created a stock of plants that produced yellow, round peas and a stock that produced green, wrinkled peas. He already knew the dominance patterns, and had already concluded that parents bore two 'unit factors' (genes) for each trait. So, he hypothesized that the genotypes of the plants in each pure-breeding stock would be RRYY (round, yellow) and rryy (wrinkled green). He placed pollen from the wrinkled, green stock on flowers of the yellow, round stock and observed the progeny.
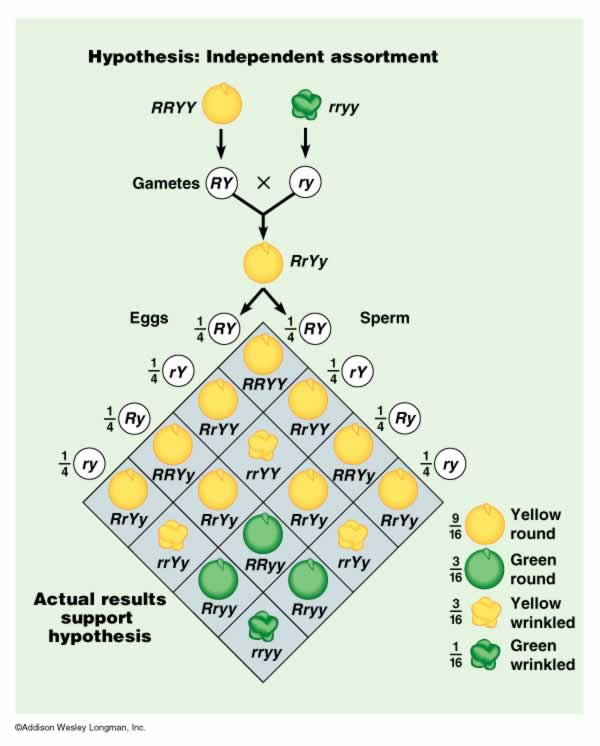 Round, Yellow (RRYY) x wrinkled, green (rryy)
Round, Yellow (RRYY) x wrinkled, green (rryy)
He hypothesized that the parental types would produce RY gametes and ry gametes, respectively. The combining of the genes in the offspring would make only RrYy genotypes, that should all express the phenotypic traits of yellow, round peas. This hypothesis was confirmed. However, this did not yet address the issue of independence; this pattern would also have occurred if color and shape were governed by one gene, or if the genes were inherited in a dependent manner. In short, the Y had to be inherited with an R, because that's all the RRYY parent had to give.
b. Crossing the F1's:
He crossed the F1 individuals, and found the following results:
- 315 round, yellow peas (~ 9/16)
- 101 wrinkled yellow peas (~3/16)
- 108 round green peas (~3/16)
- 32 wrinkled green peas (~1/16)
TOTAL - 556 peas
c. His interpretation and hypothesis:
First, it was obvious that the two traits were not caused by the same gene; all yellow peas were not ALSO round - he had produced some yellow wrinkled peas. So, different genes governed see color and seed shape, and so the genes for these traits could occur in differnt combinations.
Mendel realized that the 3:1 ratios were still preserved for each trait, when considered separately. For example, taken together, the ratio of yellow:green peas was 416:140 (2.97:1) and the ratio of round:wrinkled peas was 423:133 (3.18:1).
And also, Mendel realized that the
results of this experiment, and the results of subsequently self-crossing these
hybrids, were consistent with the probability of occurrence of independent events.
We know this as The Product Rule: "The probability that independent events
will occur together is the product of their independent frequencies."
These ratios in which the traits occurred in combination equaled the products of the independent frequencies:
3/4 W X 3/4 Y = 9/16 WY (round yellow)
1/4 w X 3/4 Y = 3/16 wY (wrinkled yellow)
3/4 W x 1/4 y = 3/16 Wy (round green)
1/4 w x 1/4 y = 1/16 wy (wrinkled green)
Mendel's hypothesis can be stated like this:
During gamete formation, the way one pair of genes (governing one trait) segregates is not affected by (is independent of) the pattern of segregation of other genes; subsequent fertilization is random. This is called Mendel's Second Principle: Principle of Independent Assortment.
He tested this hypothesis is subsequent experiments using three traits (seed color, seed shape, and seed coat color), and showed that the combination of traits expressed in the offspring were consistent with predicitons from their independent frequencies. For example, In a "trihybrid" F1 cross (a self-cross of an individual that is heterozygous for all three traits - YyRrCc), the fraction of peas that are yellow, round, and have a brown seed coat (caused by the dominant allele, "C") should be 3/4 x 3/4 x 3/4 = 27/64. His results confirmed his hypothesis.
Study Questions:
1. How did Darwin explain the existence of 'convergent communities"?
2. The Galapagos are dominated by many unique species of reptiles, birds, and marine mammals. What did this non-random assemblage suggest to Darwin about their origin, and how was evolution implied?
3. Why were the mockingbirds so critical to Darwin's ideas about the production of new species?
4. Outline Darwin's argument of the fact of evolution.
5. How did Darwin use the comparative method and observations of 'artificial selection' to produce the theory of 'natural selection'?
6. How were Malthus's observations and conclusion rlevant to the development of Darwin's theory?
7. Outline the theory of natural selection as an argument, with three premises, 3 conclusions, and a corollary.
8. How did Darwin solve Paley's dilemma regarding the stepwise evolution of a 'camera' eye?
9. How did Darwin explain the absence of LIVING intermediate forms?
10. How did Darwin explain the absence of EXTINCT intermediate forms?
11. How did Darwin believe that variation was produced in natural populations?
12. Outline Darwin's model of evolution, listing 'sources of variation' and 'causes of evolutionary change'.
13. List Mendel's four postulates, clearly and specifically stating his 'first principle' of heredity without using the word 'segregate' in your definition..
14. Why is a test cross an effective way of determining the genotype of an individual?
15. Determine the genotypic and phenotypic ratios from the following crosses. Assume dominance of capital letter:
a. AA x AA
b. AA x Aa
c. Aa x Aa
d. Aa x aa
e. aa x aa
16. Why did Mendel conduct reciprocal crosses? What hypotheses did this test?
17. Why did Mendel conduct an F1 x F1 cross (the second generation of crosses)? What hypothesis did this test?
18. How did the results from the reciprocal crosses suggest that both parents must contribute genetic information?
19. What question was Mendel addressing with his dihybrid cross?
20. How did the results lead him to hypothesize that the traits must be inherited independently? Use the product rule in your answer.
21. State Mendel's second principle, without using the word 'independent' in your definition.