 We
have summarized some of the major epochs in the history of the Earth before,
but we will review them here and add in important evolutionary events in the
history of life.
We
have summarized some of the major epochs in the history of the Earth before,
but we will review them here and add in important evolutionary events in the
history of life. The use of fossils and genetic phylogenies have allowed scientists to reconstruct the history of life and the major epochs and innovations of evolution that have occurred over the last 4 billion years of earth's history. Perhaps the most surprizing thing that we have learned is that we humans are very late arrivals on the evolutionary stage, and that the play had been going on just fine for quite a long while without us. Indeed, the evolution of anything you might recognize as an 'animal' is a fairly recent event in the evolutionary history of the planet; animals have only been present for the last 1/7th of earth history. Although we might think of macroscopic animals and plants as the 'most important' forms of life somehow, it is really the little things - bacteria and archaeans - that keep the biosphere running and allow animals and plants to survive. Without bacteria, you would not digest your food efficiently, plants would have a more difficult time absorbing nutrients, and the nutrients in dead organisms would not be recycled and made available to those of us that are still alive. The basic life processes that drive the biosphere - things like nitrogen fixation, photosynthesis, respiration and energy conversion, and decomposition - are still driven by bacteria. In the first part of this lecture, we'll see that is, indeed, how it has been for the majority of earth history.
We
have summarized some of the major epochs in the history of the Earth before,
but we will review them here and add in important evolutionary events in the
history of life.
4.5 billion years ago - Earth formed (Evidence: meteorites and moon rocks date to this age, dating the condensation of planetissimals in the solar system)
4.0 bya - oldest rocks (The oldest rocks are not as old as the Earth because the Earth's surface was molten - as a result of constant meteorite bombardment and the energy released during the stratification of Earth's core, mantle, and crust. In addition, the Earth is tectonically active, recycling its crust. So, older rocks that may have solidified may have been subducted and melted)
3.5-3.8 bya - origin of life (The oldest fossils, both microfossils and stromatolites, date to 3.5-3.8 bya). These organisms are similar to prokaryotes we see today. However, it is not possible to distinguish whether they are Bacteria or Archaeans. However, from this period forward there are anaerobic and non-oxygenic photosynthetic bacteria.
2.5-2.0 bya - oxygen accumulating ("red bed" sediments form, demonstrating that oxygen gas in the water was oxidizing iron); this is evidence of the evolution of oxygenic photosynthesis. With the accumulation of oxygen, many obligate anaearobes undoubtedly go extinct. However, other bacteria evolve the capacity to USE oxygen as an oxidizing agent, making the harvest of energy from glucose metabolism more efficient... in other words, aerobic respiration evolves.
1.7-2.0 bya - eukaryotes evolve through endosymbiotic absorption/tolerance of aerobic bacteria (protomitochondria). With the evolution of eukaryotes comes the evolution of sexual reproduction. These endosymbionts may have been a cellular parasite, like Rickettsia. Subsequent lines of eukaryotes absorbed photosynthetic cyanobacteria, which evolved into symbiotic chloroplasts. The evidence for these theories of eukaryote evolution are the genetic similarity between mitochondria and chloroplasts (that have their own DNA) and various types of bacteria and cyanobacteria. So until this point - representing nearly 1/2 of life's history on this planet - life was exclusively bacterial. The major types of ecological roles, from nitrogen fixation, to photosynthesis, to heterotrophy, to saprophagy (eating dead stuff) all evolved among bacteria - creating food webs and biogeochemical cycles that shunted energy through the biosphere and other earth systems.
0.7-0.9 bya - the first animals evolve - sponges and then soft-bodies invertebrate fossils, often radially symmetrical like jellyfish.
0.5 bya - radiation of animals with hard bodies (Cambrian explosion) - beginning of the Paleozoic Era ('old animals')
0.4 bya - evolution of land animals and plants
0.24 bya (240 mya) - Permian extinction; end of the Paleozoic Era
240-65 mya - Mesozoic Era (Age of Dinosaurs - 'middle animals')
65 mya - Cretaceous extinction (meteorite impact off Yucatan Peninsula) - beginning of the 'Cenozoic Era' ('recent animals')
6-5 mya - split of chimp and human lineages
4.5 mya - Australopithecines (1/1000 of earth's history left to present...99.9 % has occurred already).
0.2 mya (200,000) - oldest Homo sapiens fossil. (99.995 % of earth's history has passed).
II. The Prokaryote Domains: Bacteria and Archaea
A. Overview
 These
two domains are ancient; they may even have diverged before 3.5 bya. The problem
with using genetic clocks and genetic similarity to attempt to date this point
of genetic divergence on these groups is that prokaryotes exchange DNA "laterally".
They don't just pass it down a lineage from parental cell to daughter cells;
they can also pass genes from one living bacterium to another through conjugation
or transformation. As such, genetic similarity can occur as a result of lateral
gene transfer between unrelated organisms, and thus the degree of genetic similarity
in particular genes does not necessarily indicate the degree of common ancestry.
For these reasons, it is very difficult to use this particular technique to
date the time of divergence of these two ancient groups.
These
two domains are ancient; they may even have diverged before 3.5 bya. The problem
with using genetic clocks and genetic similarity to attempt to date this point
of genetic divergence on these groups is that prokaryotes exchange DNA "laterally".
They don't just pass it down a lineage from parental cell to daughter cells;
they can also pass genes from one living bacterium to another through conjugation
or transformation. As such, genetic similarity can occur as a result of lateral
gene transfer between unrelated organisms, and thus the degree of genetic similarity
in particular genes does not necessarily indicate the degree of common ancestry.
For these reasons, it is very difficult to use this particular technique to
date the time of divergence of these two ancient groups.
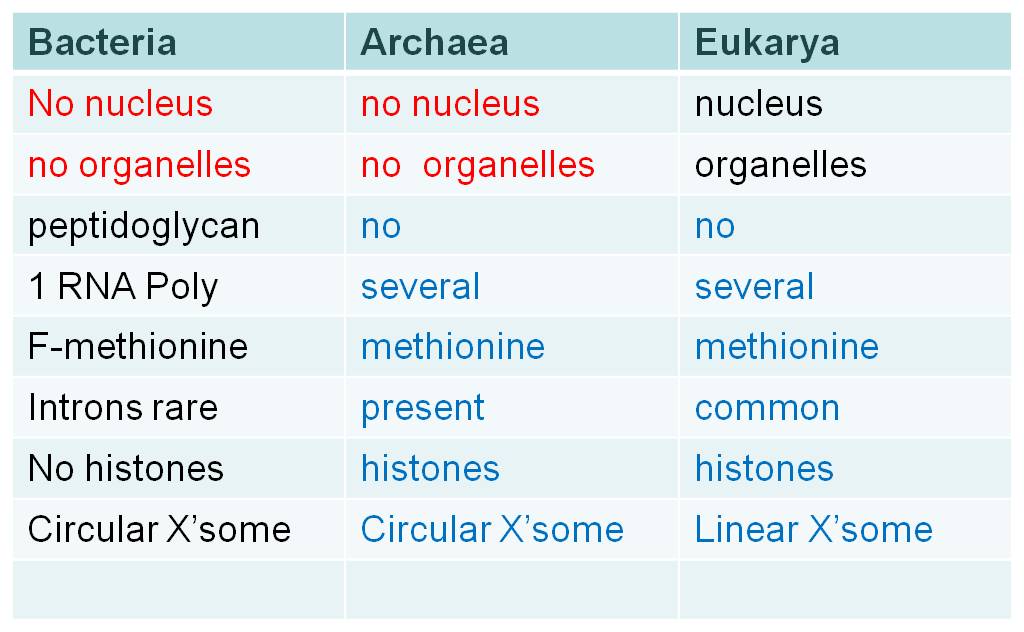
Although they share some fundamental similarities like the lack of a nucleus, the lack of membrane-bound organelles, and circular chromosomes, these two prokaryotic domains differ in some important respects. When they differ, the archaeans are often similar to the Eukarya. For example, bacteria have peptidoglycan in their cell walls, while archaeans (and eukaryotes) do not. Bacteria have only one type of RNA Polymerase; archaeans (and eukaryotes) have several. Bacteria use formyl-methionine as the initiator amino acid in translation, archaeans (and eukaryotes) use methionine. Histones are not associated with bacterial chromosomes, but they are associated with archaean (and eukaryote) chromosomes. Introns are rare in bacterial genes, but are progressively more common in archaeans and eukaryotes. So, there are several fundamental differences between these two groups, and often the archaeans are more similar to eukaryotes than to bacteria.
1. Archaea
 The
archaea are very diverse, and include groups that have adapted to some of the
most inhospitable environments on Earth. Indeed, many of these environments
may be similar to those that were common in early earth history. So, rather
than thinking that these organisms have evolved to exploit these isolated odd
habitats, perhaps they adapted to these conditions when the conditions were
common, and have just 'hung on' in these habitats that have become rare over
time as the earth has changed. The extreme thermophiles
live at very high temperatures at sulphur springs and geothermal vents where
temperatures can exceed 1000C. How they protect their proteins and
DNA under these conditions is an area of active research. Another group are
tolerant of very extreme salt concentrations - these are the extreme halophiles.
Yet another group of extreme organisms are the methanogens. These organisms
use H2 to reduce CO2 and produce energy; producing CH4
(methane or 'swamp gas') as a waste product. Most are obligate anaerobes, meaning
that they can only survive in environments where the concentration of oxygen
gas is very low. However, some are aerotolerant, and can survive in the presence
of oxygen though they do not use it for respiration. These conditions occur
at the bottom of deep water bodies, or where the action of respiring bacteria
deplete available oxygen - like in swamp and marsh sediments and in the digestive
tract of animals (including humans). Some methanogens are extreme thermophiles,
too.
The
archaea are very diverse, and include groups that have adapted to some of the
most inhospitable environments on Earth. Indeed, many of these environments
may be similar to those that were common in early earth history. So, rather
than thinking that these organisms have evolved to exploit these isolated odd
habitats, perhaps they adapted to these conditions when the conditions were
common, and have just 'hung on' in these habitats that have become rare over
time as the earth has changed. The extreme thermophiles
live at very high temperatures at sulphur springs and geothermal vents where
temperatures can exceed 1000C. How they protect their proteins and
DNA under these conditions is an area of active research. Another group are
tolerant of very extreme salt concentrations - these are the extreme halophiles.
Yet another group of extreme organisms are the methanogens. These organisms
use H2 to reduce CO2 and produce energy; producing CH4
(methane or 'swamp gas') as a waste product. Most are obligate anaerobes, meaning
that they can only survive in environments where the concentration of oxygen
gas is very low. However, some are aerotolerant, and can survive in the presence
of oxygen though they do not use it for respiration. These conditions occur
at the bottom of deep water bodies, or where the action of respiring bacteria
deplete available oxygen - like in swamp and marsh sediments and in the digestive
tract of animals (including humans). Some methanogens are extreme thermophiles,
too.
Although the Archaea are noted for representatives that tolerate these extreme environments, there are other archaeans that live under more benign environmental conditions across our planet, where they function as photosynthetic producers, consumers, and decomposers. Curiously, although archaeans do inhabit the human gut, no archaeans have been identified that cause human disease. The other prokaryotes - the bacteria - contain many species that cause human disease.
2. Bacteria
 The
bacteria are as diverse than the archaeans, expressing an extraordinary variety
of metabolic processes. A couple notable groups are the cyanobacteria, thought
to share common ancestry with eukaryotic chloroplasts, and the gram-positive
bacteria that have many types of pathogenic bacteria, such as those causing
staph and strep infections, botulism, tuberculosis, anthrax, and many other
diseases.
The
bacteria are as diverse than the archaeans, expressing an extraordinary variety
of metabolic processes. A couple notable groups are the cyanobacteria, thought
to share common ancestry with eukaryotic chloroplasts, and the gram-positive
bacteria that have many types of pathogenic bacteria, such as those causing
staph and strep infections, botulism, tuberculosis, anthrax, and many other
diseases.
Both the bacteria and archaea are very old groups, and have been radiating and evolving for over 3.0 billion years. Recent genetic studies have revealed an astounding degree of genetic variation within these groups - an understandable consequence of their long evolutionary histories. This genetic variation is reflected by the 'branch lengths' in the tree diagram to the right. The genetic variation among the plants, animals, and fungi, represented by their relative short branch lengths, are dwarfed by the genetic variation among the archean and bacterial clades. But of course, the animals have only been radiating for the last 0.7 billion years. So, the archean and bacterial lineages have been radiating for 5 times as long and so it is not surprizing that they are much more diverse. Thus, as expected for groups that are so old, they have radiated dramatically - even within subgroups. For this reason, even groups that share common ancestry can include organisms that are very different. For our purposes here, an overview of the diversity is presented, rather than a overview of their classification or phylogeny.
If we examine the morphological diversity in prokaryotes, we find round (coccus), rod-shaped (bacillus), and spiral (spirochaete). That is not a lot of morphological diversity! So how have the prokaryotes been changing over the last 3.5 by? Well, they have been evolving metabolically. The theme of bacteria diversity is metabolic diversity. There are very few metabolic processes that eukaryotes can perform that prokaryotes cannot. However, the reverse is not true; the prokayotes have a wide range of metabolic diversity - a range that, in many respects, exceeds the metabolic diversity of eukaryotes.
1. Requirements of Oxygen for Cellular Respiration
Consider types of respiration. There
are no anaerobic eukaryotes. Most eukaryotes require oxygen (they have mitochondria,
after all), although some can function without it for a short time (facultative
anaerobes like yeast). Prokaryotes, on the other hand, have a wide range of
'respiratory' strategies:
- some bacteria, like methanogens, cannot live in the presence of oxygen - obligate
anaerobes
- some bacteria can tolerate oxygen
but don't use it - aerotolerant
- some bacteria can live with or without oxygen - facultative aerobes
- some bacteria live only in the presence of oxygen - obilgate aerobes
2. Metabolic Pathways for Harvesting Energy and Carbon from the Environment
1. Chemolithotrophs:
Use H2S or NH3 as an electron donor to power ATP production without sunlight;
and then use ATP to synthesize glucose from CO2. So, they harvest their
own energy from chemicals and make their own large organic molecules.
This is only done by bacteria.
2. Photoautotrophs:
Use either H2S (non-oxygenic) or H2O (oxygenic) photosynthesis powered by sunlight
to make ATP. Use the ATP to make glucose. So, they harvest energy
from light and make their own carbon resource. Cyanobacteria are oxygenic, and
their ancestors are probably responsible for the oxygen revolution about 2.5-2.0
bya. In addition to bacteria, some protists (algae) and most plants use this
metabolic strategy. The genetic similarity between chloroplasts (in algae and
plants) and certain cyanobacteria suggest that cyanobacteria may be ancestral
to chloroplasts,having been engulfed by phagocyctosis billions of years ago.
3. Photoheterotrophs: These strange organisms use sunlight as a source of energy for the formation of ATP, but they do not synthesize their own large carbon molecules (glucose). Rather, they have to absorb these molecules from the environment (heterotrophic). This is only done by some types of bacteria.
4. Chemoheterotrophs - must get BOTH energy and carbon from the metabolism of complex organic molecules. This is the most common strategy for living systems, exploited by some bacteria, some protists, all fungi, and all animals.
3. Ecological Roles
- primary producers: bacteria still make a significant contribution to the absorption of CO2 and the production of oxygen on our planet - largely in the oceans. And these oceanic bacteria are the bottom of marine food chains.
- nitrogen-fixing bacteria: can convert N2 into NH3 (ammonia) and then NO3 (nitrates) which can be taken up by plants. Only bacteria can do this; and so all new, natural inputs of nitrogen to the biosphere are by bacteria. (Lots of biologically accessible nitrogen is added to the environment by humans - through fertilizer and the production of NOx compounds by internal combustion engines).
- decomposers: as consumers of dead organic matter, bacteria (and fungi) break down complex organics and harvest energy. And the byproducts of that digestion are mineral nutrients (N, P, K) that are made available for reuptake in the biosphere by plants. So, bacteria are needed for nutrients to cycle through ecosystems.
- pathogenic bacteria: these bacteria get energy from living tissue and destroy cells in the process; they also produce either exotoxins that they secrete, or endotoxins that are bound to the cell wall. Examples of these pathogenic bacterial diseases are: staph infections, salmonella, typhoid, cholera, scarlet fever, tetanus, tuberculosis, botulism, gonorrhea, and syphilis.
- symbiotes: there are gut bacteria in nearly all animals that assist with digestion and may be required for the production and absorption of certain metabolites.There are also bacterial symbionts in plants, protists, and fungi.
As such, bacteria interact in every way possible with other organisms, and form the 'hub' of the biosphere. In some ways, the evolution of life has been an "embellishment" of the bacterial ecosystems that dominated the planet for the first 2.3 billion years of life.
The take home message about prokaryotes is their metabolic diversity. For 1.8 billion years, life on Earth was SOLELY bacterial. How was it evolving? How was it responding to the environment? Well, it seems that life was responding metabolically, adapting by exploiting different environments and energy sources in different ways.
The eukaryotes have been evolving for at least 2.0 billion years. As you might expect (as was argued for the prokaryotes groups), there has been considerable divergence within this group, producing a dramatic range of genetic and metabolic variation. And of course, the multicellular eukaryotes evolved from protistan ancestors - so multicellular eukaryotes (plants, fungi, and animals) are included within this clade. So, the focus will be on the origin of the clade (Eukarya) and the diversity within this clade. We will pay particular attention to the sister groups of the multicellular "kingdoms" that we will consider in greater detail in the remaining lectures. Obviously, the features that distinguish eukaryotes from archaean ancestors are the presence of membrane-bound organelles (mitochondria and chloroplasts) and a nucleus. So, any hypothesis of origin must explain how these defining structures came into existence.
1. Origin of Organelles
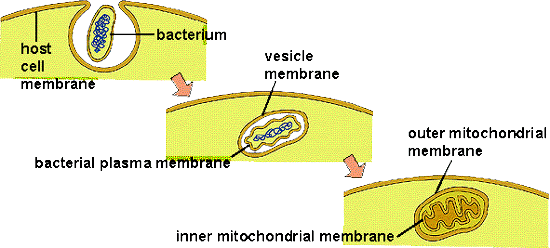
The origin of eukaryotic cells is described by the Endosymbiotic Theory, popularized by Lynn Margulis in the 1960's. It states that organelles are descended from free-living bacterial cells that were absorbed by - or parasitized - archaeans. The evidence for this idea is very strong. Both mitochondria and chloroplasts have double membranes. The outer membranes are similar to the plasma membrane of eukaryotes and archaeans. The inner membrane has certain attributes of bacteria. If a bacterium were absorbed through the process of endocytosis, the bacterium would be surrounded by the membrane of the host's vacuole - creating a double membrane. In addition, both types of organelles have their own DNA, which is circular like bacterial DNA. Both mitochondria and chloroplasts divide by fission, independent of the mitotic division of the eukaryotic cell they inhabit. Over the course of evolutionary history, genes have been exchanged between the organelles and the nucleus, or have been eliminated in the organelle because they also exist in the nucleus. For these reasons, the organelles are functionally dependent on the eukaryotic cell and can no longer live independently for very long.
Additional similarities link these organelles to specific bacteria. Mitochondria not only have their own DNA, but they also produce their own proteins using these DNA recipes. (That should be no surprize!) Protein synthesis occurs on ribosomes within the mitochondria, and their ribosomes are more similar to bacterial ribosomes than to the ribosomes in the cytoplasm of their eukaryotic host cells (produced by the nuclear DNA). Likewise, the genetic code used by mitochondria has a few differences from the universal genetic code; differences also seen in the proteobacteria. Genetic analyses suggest that mitochondria are most closely related to Rickettsia proteobacteria. Rickettsia does not causes "rickets" (a vitamin-D deficiency); rather, Rickettsia bacteria cause a variety of human diseases like many forms of tick-borne fevers (Rocky Mountain spotted fever) and typhus. In addition to having a similar genome to mitochondria, these bacteria are obligate intracellular parasites - so they move into eukaryotic cells and become "organelle-like". In addition, Rickettsia produce ATP in the same way that mitochondria do. So, it is possible that archaeans absorbed bacteria and used them for ATP production, or that the bacteria infected the archaeans to consume their sugars, or some combination of these scenarios. In any case, the bacteria and archaean ultimately benefitted from the interaction, forming a mutualistic or "symbiotic" relationship.
 |
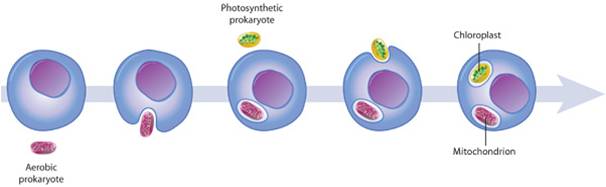 |
Chloroplasts have an independent, endosymbiotic origin; they are genetically similar to a differnt group of bacteria called cyanobacteria. Some have a double membrane like mitochondria. However, in some protistan groups, the chloroplasts have 3 or 4 membranes, suggesting multiple rounds of endosymbiosis. So, the current model of chloroplast ancestry suggests a single origin of photosynthetic red and green algae. Green algae were then absorbed (secondary endosymbiosis) by other unrelated protists, creating the euglenids and chlorarachniophytes. These organisms are not related directly through nuclear genetic analysis, but they have very similar, multi-membraned chloroplasts - suggesting independent endocytosis of green algae. A similar process may explain the chloroplast similarity in the stramenopiles, apicomplexans and dinoflagellates - independent endocytosis of a red algae could explain their dissimilar nuclear DNA and their similar, multi-membraned chloroplasts.

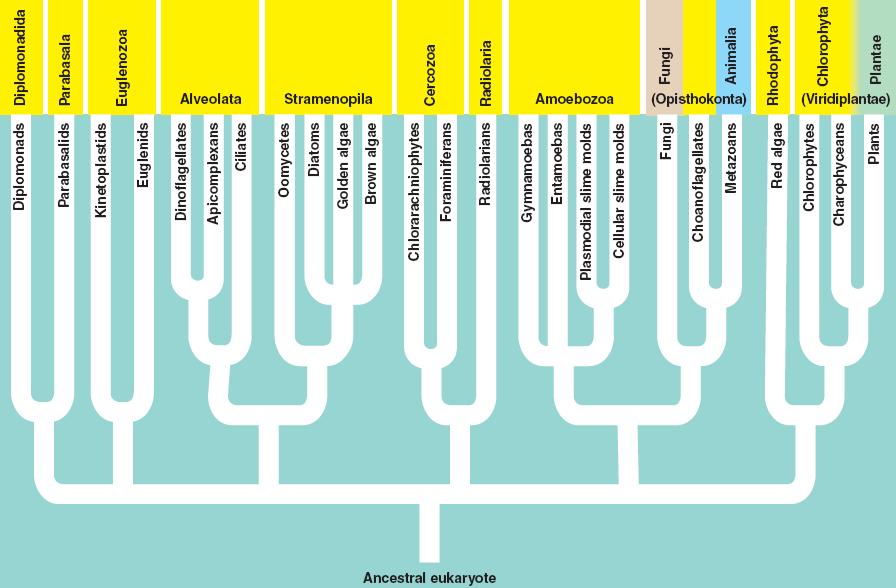
There are several groups of eukarya; most of them contain only single-celled representatives.
 7.
Opisthokonta
7.
Opisthokonta
This clade includes the fungi and animals, and the flagellated sister taxa that bear the stamp of their close phylogenetic relatedness. Other groups have flagella, like the diplomonads and the euglenozoa - but those groups have two flagella and they are at the 'front' of the organism, pulling the organism along surfaces or through a liquid medium. The Opisthokonts, however, have a single, posterior flagellum at some point in the life cycle of the organism. In the animals, this is represented by sperm cells. most fungi do not have flagellated cells, except for the most promitive group, the chytrids. This morphological characteristic is the only morphological trait that links these groups, but their genetic similarity testifies to their common ancestry.
An important opisthokont group besides the fungi and animals are the Choanoflagellates. This group is considered to be the "sister group" to the animals, and may represent what the ancestral animals evolved from. The Choanoflagellates are distinguished by a ring of small projections that ring the flagellum like a collar. These microvilli act like a sieve. The motion of the flagellum wafts bacteria into the ring of microvilli, where bacteria are trapped and then absorbed by phagocytosis. There are over 100 species of choanoflagellates; some are solitary, but some form colonies. Their predation of bacteria provides a valuable link in marine food webs. As we will see when we get to animals, the most primitive animals - the sponges - have cells that are very similar to these choanoflagellates.
 8.
Archaeplastida
8.
Archaeplastida
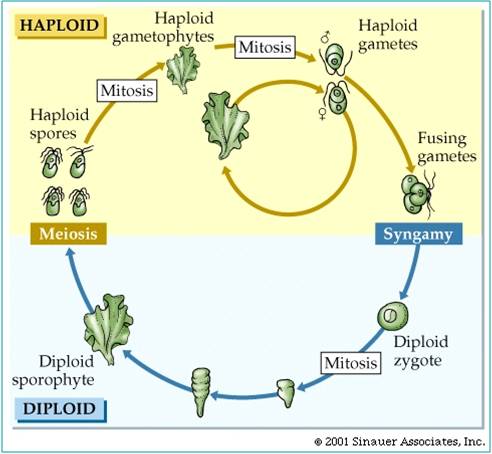
The last eukaryotes clade contains red algae, green algae, and land plants. Red algae have chloroplasts that contain chlorophyll a like cyanobacteria; green algae and plants contain chloroplasts with chlorophyll a and chlorophyll b. The red algae are largely multicellular, colonial organisms best represented by red seaweeds. Green algae are a diverse group, including unicellular, colonial, and multicellular species like Ulva and Enteromorpha (green seaweeds), Volvox, and Spirogyra. A key shared derived character that is seen in the Archaeplastida is "alternation of generation". Consider a haploid organism about to reproduce. This organism produces gametes (egg and/or sperm) by mitosis. These gametes fuse to form a diploid zygote, which developes into the diploid offspring. The diploid organism produces haploid spores by meiosis. These spores do not fertilize one another. Rather, they each divide by mitosis and develop into separate haploid organisms. So, the ploidy level alternates each generation. All plants share this life cycle, so we will see it often during the plant diversity lectures.
1. Overview:
The Opisthokonts are the eukaryote clade that includes organisms with a single posterior flagellum at some part of their life cycle. This includes the fungi, choanoflagellates, and animals. Only the most primitive group of fungi - the chytrid fungi - still have a flagellated life history stage today. Other fungi have evolved so that the spore is the dispersive life history stage.
 2.
"Body Plan"
2.
"Body Plan"
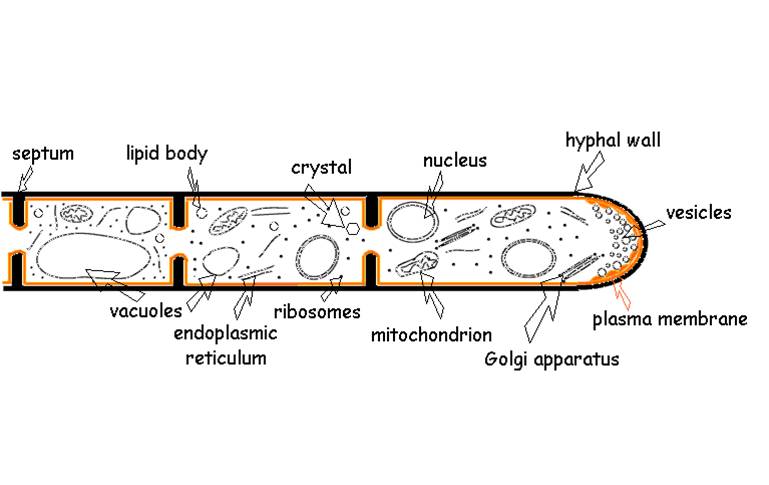
Fungi are very diverse, with over 100,000 named species. Although some fungi are unicellular (the "yeasts" - which are a diverse group taxonomically but are lumped together under this name because of their unicellular structure), most are multicellular - but with an interesting twist. The 'multicellular' fungi have a body composed of thin threads called hyphae. Hyphae have a cell wall composed of chitin - the same polysaccharide in arthropod exoskeltons. But in most species, the hyphae are not composed of discrete cells. Rather, the hyphal strand consists of multinucleate cytoplasm. In some species, there are "cell walls" called septa that subdivided the hyphal strand, but they have holes large enough to exchange mitochondria and cytoplasm. Other fungi don't even have septa. So, the cytoplasm 'streams' quickly along the length of the hyphal organism. Therefore, nutrients absorbed by one part of the fungus can be transmitted quickly through the organism. Fungi can grow very rapidly, literally growing towards nutrients concentrated in the soil or benthos. The earliest fossils that have fungal-like characteristics date to 1.4 bya, suggesting that the fungi diverged from the other opisthokont lineages early in eukaryote evolution. These fossils have septa and form a network of filaments - characteristics of many fungi we see today.
 3.
Ecological Roles
3.
Ecological Roles
Fungi are absorptive heterotrophs; they secrete enzymes into the immediate environment of the hyphae and absorb the products of that extracellular digestion. Fungi are a diverse group that play many important and interesting roles in the environment. Many digest dead material - these are the decomposers. Fungi are the primary decomposing organisms in almost all environments on earth, including marine, freshwater, and terrestrial habitats. However, as obligate aerobes, they do not perform well in anaerobic environments - in those places, anaerobic bacteria are the only decomposing organisms.
Other fungi digest living material - these are the parasitic fungi like those that cause athlete's foot, ringworm, and vaginal yeast infections (Candida albicans). Other organisms are infected by fungi, of course; indeed, fungal infections of plants called rusts, smuts, mildews, and ergots can kill the host plant and are important pathogens of crop plants. Humans and other animals that consume seeds or tissue infected with ergots can suffer hallucinations, siezures, miscarriage, and death. Chytrid fungi are significant pathogens of amphibians. Amphibian populations are crashing worldwide, probably due to a combination of global warming, habitat degradation, pollution, and the spread of chytrid fungus. Another interesting group of fungi are the 'entomophagous' fungi. Spores land on insects and germinate - boring a hole through the exoskeleton. Once inside, they digest the animal and complete their life cycle, producing spore-bearing bodies that erupt through the insects dead body. Interestingly, seelction has favored fungi that attack the nervous system last, and in a particular way - making the insects more positively phototaxic. The insects climb towards light and then die - placing the fungus above the ground where spore dispersal is more effective. There are also predatory fungi, like Arthrobotrys anchonia. This soil fungus has hyphal 'lariats'. When soil nematodes wriggle through the loop, the hyphae shunt water to the loop and it swells quickly- clamping down on the nematode. The worm is then digested. Given the extraordinary abundance of nematode worms in the soil (several thousand per cubic cm), they are a ready foodstuff for anything that can catch them - even fungi.
 Other
important fungi enter into mutualistic relationships with alga and plants. Lichens
are symbiotic relationships between algae or cyanobactera and particular species
of fungi; the photosynthetic algae or cyanobacteria provides carbon and energy
(in the form of sugars) to the fungus through photosynthesis, and the fungus
absorbs water and mineral nutrients (N and P) required by the algae. Similar
symbiosis occur between vascular plants and a particular group of fungi, the
Glomerales (or Glomales). This clade of fungi are root symbionts of plants,
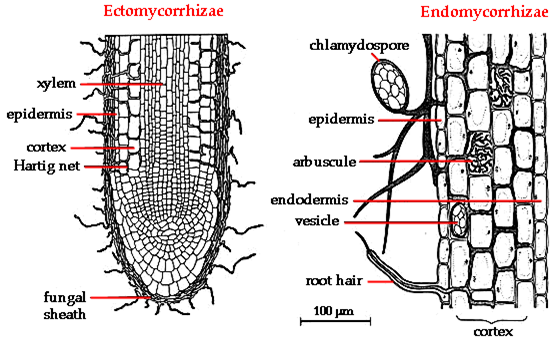
forming symbiotic relationships called mycorrhizae. There are two types of associations.
In ectomycorrhizal associations, the fungus covers the fine roots and root hairs
of a plant like a shealth, with only minor incursions of fungal hyphae into
the root, itself. In the arbuscular mycorrhizal fungi, the fungi grow specific
structures within the root, forming a much more intimate relationship. Because
hyphae are so small, they have a huge absorptive surface area per unit volume;
much greater than bulkier plant roots. And, by secreting enzymes into the soil,
they can mobilize nitrogen and phosphorus. Finally, because they grow rapidly,
they can quickly colonize areas where nutrients are available. Hyphae absorb
water and mineral nutrients and pass them to their symbiotic plant partner in
exchange for sugars that contain the energy and carbon that the fungus needs
to thrive. Over 70% of all vascular plant families have species that form mycorrhizal
symbioses with fungi. Apparently, this is a very ancient relationship, dating
back to the evolution of vascular plants. The oldest vascular plant fossils
(from the Rhynie chert - 420 mya) have fungal endosymbionts. It may be that
the evolution of mycorrhizae allowed plant to colonize land in the first place.
Other
important fungi enter into mutualistic relationships with alga and plants. Lichens
are symbiotic relationships between algae or cyanobactera and particular species
of fungi; the photosynthetic algae or cyanobacteria provides carbon and energy
(in the form of sugars) to the fungus through photosynthesis, and the fungus
absorbs water and mineral nutrients (N and P) required by the algae. Similar
symbiosis occur between vascular plants and a particular group of fungi, the
Glomerales (or Glomales). This clade of fungi are root symbionts of plants,
forming symbiotic relationships called mycorrhizae. There are two types of associations.
In ectomycorrhizal associations, the fungus covers the fine roots and root hairs
of a plant like a shealth, with only minor incursions of fungal hyphae into
the root, itself. In the arbuscular mycorrhizal fungi, the fungi grow specific
structures within the root, forming a much more intimate relationship. Because
hyphae are so small, they have a huge absorptive surface area per unit volume;
much greater than bulkier plant roots. And, by secreting enzymes into the soil,
they can mobilize nitrogen and phosphorus. Finally, because they grow rapidly,
they can quickly colonize areas where nutrients are available. Hyphae absorb
water and mineral nutrients and pass them to their symbiotic plant partner in
exchange for sugars that contain the energy and carbon that the fungus needs
to thrive. Over 70% of all vascular plant families have species that form mycorrhizal
symbioses with fungi. Apparently, this is a very ancient relationship, dating
back to the evolution of vascular plants. The oldest vascular plant fossils
(from the Rhynie chert - 420 mya) have fungal endosymbionts. It may be that
the evolution of mycorrhizae allowed plant to colonize land in the first place.
 4.
Fungal Life Cycle
4.
Fungal Life Cycle
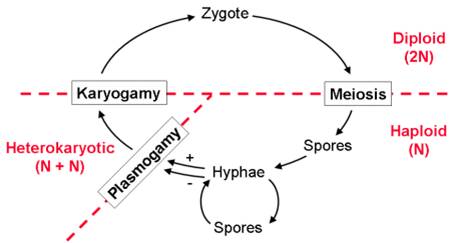
Fungi have a distinctive life cycle that includes an unusual 'dikaryotic' or 'heterokaryotic' cell type that has two nuclei. The life cycle begins when a haploid spore germinates, dividing mitotically to form a 'multicellular' haploid organism (hypha). In most fungi, hyphae can produce spores directly, through an asexual process (mitosis). Hyphae belong to different sexual types, represented by their ability to "mate". When different mating types contact one another, the cytoplasm of the hyphae fuse but the nuclei remain separate. This process is called 'plasmogamy', and it creates an initial cell called a 'dikaryon'. If it divides to produce multicellular tissue, both nuclei divide and are passed on to both daughter cells - creating dikaryotic ( 'heterokaryotic' in the figure) tissue. Eventually the nuclei fuse, creating a true diploid cell ("zygote"). This cell undergoes meiosis, producing haploid spores that are dispersed to the environment.
 1.
Green Algal Ancestors
1.
Green Algal Ancestors
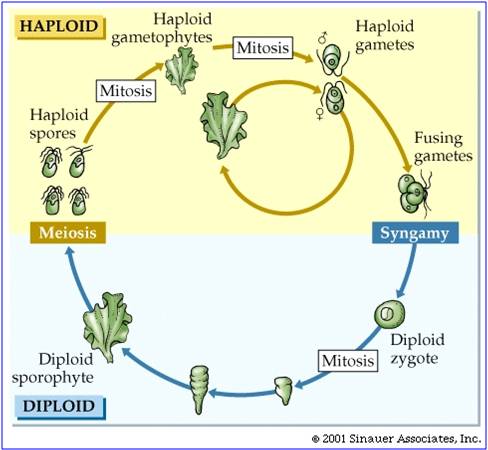
There is considerable evidence that plants evolved from green algal ancestors. They share the same type of chloroplasts (with chlorophyll a and b), and an intersting life history called "alternation of generation". In this life cycle, a haploid organism (the gametophyte) produces gametes by mitosis. These gametes fuse in a fertilization event, creating a diploid zygote. This diploid zygote divides (typically) mitotically to produce a multicellular diploid offspring. This organism produces haploid spores by meiosis. These spores develop, dividing mitotically to produce the next multicellular haploid generation. So, the ploidy level "alternates" across the generations. In green algae, the sporophyte and gametophyte stages may be "isomorphic" (look the same), like in sea lettuce (Ulva) show at right. In addition, green algae exhibit a range of gamete types, from isogametes that are double flagellated and the same size (as in Ulva) to oogamous species with double-flagellated sperm and immobile large eggs. All plants are oogamous, with large eggs and small sperm. In some gymnosperms and the angiosperms, the sperm have no flagella; in all other plants the sperm have flagella.
2. Colonization of Land: Environmental Differences
 The evolution of plants occurred as photosynthetic organisms acquired traits
that allowed for greater reproductive success on land - out of the ancestral
aquatic environment of algae. So, to understand how plants differ from algae,
and how and why plants evolved various characteristics, you must understand
how aquatic and terrestrial habitats differ. Algae and plants require several
essential nutrients: sunlight, water, and mineral nutrients (like nitrogen,
potassium, magnesium, and phosphorus, to name a few). They also require CO2
and oxygen, as all green algae and plants are photosynthetic aerobes, having
both chloroplasts (for photosynthesis that needs CO2) and mitochondria
(for aerobic respiration that needs O2). Aquatic habitats have ample
water (obviously). Carbon dioxide dissolves in water, so the concentration in
air and water will equilibrate over time. However, CO2 does not diffuse
quickly into or through water, so it may be less abundant locally. Likewise,
oxygen may be in short supply in aqueous habitats, largely because it does not
dissolve as readily. Although oxygen has a concentration of 21% in the atmosphere,
it is often about 7% or less in aquatic habitats, with more near the surface
and less at increasing water depths. In addition, water absorbs light energy
quickly. About 90% of the light energy is absorbed in the first 10m of water;
99% is absorbed by 50-100m. So, photosynthetic organisms must stay near the
top of a water column to get light and oxygen. Unfortunately, the process of
decomposition that recycles mineral nutrients tends to occur at the bottom of
a water column. As such, growth and photosynthesis of aquatic algae is usually
strongly limited by mineral nutrients - either phosphorus, nitrogen, or iron.
(This is why fertilizers running of into water can create huge algal blooms
- the factor previously limiting their growth has been provided, so they grow
like crazy). That is why the open ocean has little photosynthetic activity per
unit area; especially compared to shallower regions like continental margins
and shallow estuaries where upwelling and wave action bring nutrients to the
surface and feed local productivity. So, although aquatic habitats have lots
of water, terrestrial environments provide more sunlight, more oxygen, the same
amount of CO2 (but more equitably distributed and available), and
more mineral nutrients (which are eroding from the rocks or are being recycled
by decomposers and are present in the soil water). And in shallow-water environments
with a supply of nutrients, competition with other algae would become intense,
too.
The evolution of plants occurred as photosynthetic organisms acquired traits
that allowed for greater reproductive success on land - out of the ancestral
aquatic environment of algae. So, to understand how plants differ from algae,
and how and why plants evolved various characteristics, you must understand
how aquatic and terrestrial habitats differ. Algae and plants require several
essential nutrients: sunlight, water, and mineral nutrients (like nitrogen,
potassium, magnesium, and phosphorus, to name a few). They also require CO2
and oxygen, as all green algae and plants are photosynthetic aerobes, having
both chloroplasts (for photosynthesis that needs CO2) and mitochondria
(for aerobic respiration that needs O2). Aquatic habitats have ample
water (obviously). Carbon dioxide dissolves in water, so the concentration in
air and water will equilibrate over time. However, CO2 does not diffuse
quickly into or through water, so it may be less abundant locally. Likewise,
oxygen may be in short supply in aqueous habitats, largely because it does not
dissolve as readily. Although oxygen has a concentration of 21% in the atmosphere,
it is often about 7% or less in aquatic habitats, with more near the surface
and less at increasing water depths. In addition, water absorbs light energy
quickly. About 90% of the light energy is absorbed in the first 10m of water;
99% is absorbed by 50-100m. So, photosynthetic organisms must stay near the
top of a water column to get light and oxygen. Unfortunately, the process of
decomposition that recycles mineral nutrients tends to occur at the bottom of
a water column. As such, growth and photosynthesis of aquatic algae is usually
strongly limited by mineral nutrients - either phosphorus, nitrogen, or iron.
(This is why fertilizers running of into water can create huge algal blooms
- the factor previously limiting their growth has been provided, so they grow
like crazy). That is why the open ocean has little photosynthetic activity per
unit area; especially compared to shallower regions like continental margins
and shallow estuaries where upwelling and wave action bring nutrients to the
surface and feed local productivity. So, although aquatic habitats have lots
of water, terrestrial environments provide more sunlight, more oxygen, the same
amount of CO2 (but more equitably distributed and available), and
more mineral nutrients (which are eroding from the rocks or are being recycled
by decomposers and are present in the soil water). And in shallow-water environments
with a supply of nutrients, competition with other algae would become intense,
too.
In this context, land looks like a much better habitat for photosynthetic organisms than aquatic habitats. We can appreciate why selection might favor organisms that could colonize this environment. However, there is one significant hurdle - terrestrial environments are very very dry. Living tissue is mostly water; in order to survive in a terrestrial environment, living systems had to evolve mechanisms for reducing water loss. There are other problems that a deficit of water creates, too, like for reproduction. Algae and animals have swimming gametes; how can they reproduce on land? As we consider different groups of plants, we will focus on how plants evolved characteristics that solved these problems.
 3.
Plant Evolution: Adapting to Land
3.
Plant Evolution: Adapting to Land
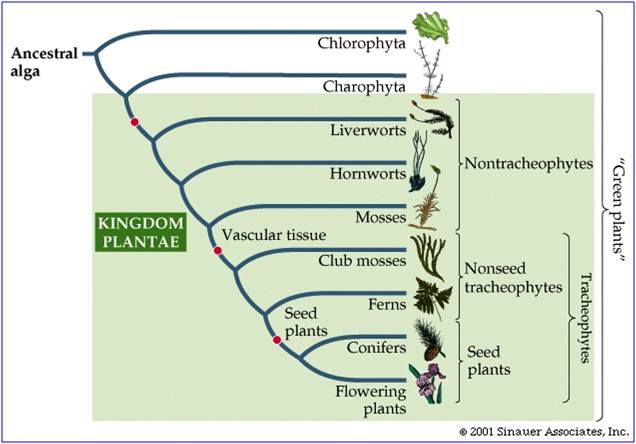
The dominant plants on the planet today are the flowering plants (Angiosperms) and the conifers (a group of Gymnosperms). Both groups are fully terrestrial (except those that have recolonized water!), meaning that no stage of their life cycle requires an aquatic habitat to survive. They have a waxy cuticle on their leaves that reduces water loss, and they also have pores ("stomates") that close during dry conditions to reduce water loss still further. They have vascular tissue for the efficient transport of water from the roots to leaves ("xylem" tissue) and to transport sugars from leaves to the rest of the plant ("phloem" tissue). The xylem has cellulose and lignin in the cell walls, creating rigid tissue that can support the weight of the plant. In addition, these plants have non-motile sperm; the sperm is carried to the egg by the male gametophyte organism, the pollen grain. All of these characteristics are adaptations to life on land: reducing water loss, increasing the efficiency of water transport, and reducing dependancy on water as a necessary medium for gamete transport or bodily support. These adaptations did not evolve at once. Rather, through the evolution of different new plant groups, plants evolved and accumulated these adaptations. Today, although angiosperms dominate the plant kingdom, there are still representatives of more ancient groups. As we move through these groups, we will emphasize the major evolutionary innovations of each group.
4. Primitive Plants - the "non-tracheophytes"
 a.
Groups
a.
Groups
These groups are not a 'monophyletic' clade, but we will consider them together. They are the "liverworts", the "hornworts", and the "mosses". You are certainly aware of mosses, but liverworts are quite common, too, living on streambanks or on rocks in streams. Hornworts are a bit less common, but are also found in moist areas, including the bark of some trees.
b. Characteristics
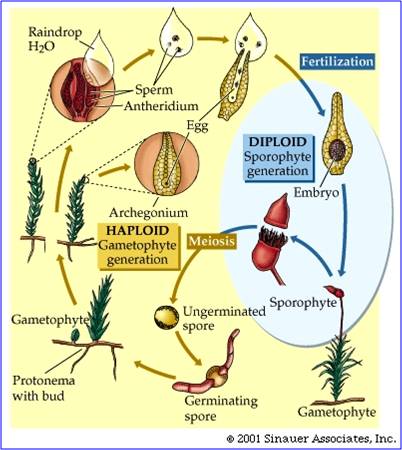
These groups are all non-vascular, meaning that they do not have xylem or phloem. As a consequence, water and sugar transport in the plants is from cell to cell and it is not very efficient. So, the plants must remain small and low to the ground - the source of their water. They also produce swimming sperm, and so must be covered by a thin film of water in order to reproduce. They do not have true leaves or roots (defined as having veins), and so water absorption is inefficient, too. In short, these plants are limited to living in very moist areas, and they must be covered by at least a thin film of water at some time during the year. These plants do have some adaptations for life on land that distinguish them from their algal ancestors, however. They secrete a thin oily cuticle that protects them a bit from water loss. In addition, hornworts and moss sporophytes have stomates - pores in their 'leaflets' to regulate gas exchange and water loss. Their life cycle is dominated by the haploid gametophyte generation; that means that when you are looking at a green moss plant or a liverwort, you are looking at a haploid gametophyte organism. The sporophyte organism is smaller, and is physiologically dependent on the haploid gametophyte parent. In moss, the sporophyte grows out of the archogonium (egg-bearing structure) on the top of the female moss gametophyte. It is brown, and receives the energy it needs to grow from the parent plant. The capsule that it produces is where meiosis occurs, reducing ploidy by 1/2 and producing the haploid spores that will each develop into new haploid gametophyte plants.
c. History
The evolutionary history of these plants is scant, because they do not produce thick, lignified cell walls that fossilize. However, scientists working in Argentina unearthed what they believe to be Liverwort spores that date to approximately 470 mya. This is the earliest fossil record of a terrestrial plant group.
5. Tracheophytes
 a.
Characteristics
a.
Characteristics
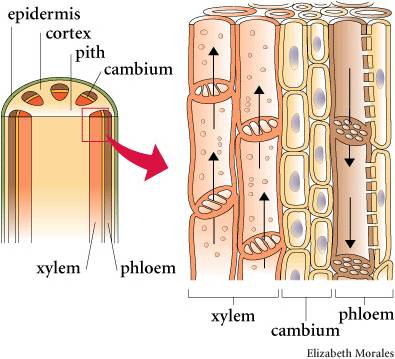
As the name denotes, these are the vascular plants that have true xylem and phloem tissue. Xylem tissue consists of cells that have died, leaving behind their highly lignified cell walls. The ends of these vessel elements have holes, allowing water to pass through like a straw. The evolution of xylem allows for efficient transport of water up a plant. This occurs through root pressure and evapotranspiration from leaves. By actively transporting salts into the xylem cavity, water follows by osmosis from living root tissue and water pressure 'pumps' water up the stem. At the other end of the xylem vessels, water is evaporating off leaves to the environment, creating a negative pressure that 'sucks' water up the stem. Because of the narrowness of the vessel elements, water also moves up by capillary action. All this means that plants could grow taller. This is advantageous for gathering light (the plant is not subject to shading by neighbors) and dispersing spores, but not too great for dispersing swimming sperm.
As such, it is not surprizing that the sporophyte generations becomes the dominant life history stage in tracheophytes. The gametophyte can't take advantage of larger size, because the plant needs to be covered by water for the sperm to swim. The sporophyte, however, benefits from being tall because the increased wind disperses spores farther.
b. Groups:
i. Non-seed tracheophytes: These are the most primitive living vascular plants, descended from a very diverse clade that dominated through the late Silurian, Devonian, and Carboniferous Periods (400-300 mya) of the Paleozoic Era. The presence of vascular tissue allowed their sporophytes to grow tall; indeed, the 'arborescent' (tree-like) canopy of Devonian and Carboniferous forests were clubmosses nearly 100 ft tall. Today, clubmosses are typically under a foot in height, forming a lush carpet in the herb layer of moist forests. The life cycle shows this inversion and dominance of the sporophyte. When you are looking at a green clubmoss plant, you are seeing the diploid sporophyte. It bears cone-like structures called strobili, and in those structures the sporangia produce haploid spores by meiosis. The spores germinate, producing a free-living gametophyte that lives in the soil. Archegonia and antheridia produce eggs and sperm, respectively, by mitosis. The swimming sperm fertilize the eggs in the archegonia, and the developing sporophyte grows out of its gametophyte parent, as in the more primitive true mosses. However, this sporophyte is photosynthetically active and grows to be a large, physiologically independent organism - it is not dependent on its gametophyte parent as in true mosses.
 The
fern life cycle is very similar to the clubmosses; indeed, these groups were
sometimes grouped together. Ferns have a dominant sporophyte that produces true
leaves (fronds). In many species, the leaves bear sporangia underneath, which
produce spores by meiosis. The haploid spores are released, and they develope
into small, independent gametophytes that live in the leaf litter. The gametophytes
bear antheridia and archegonia, that produce sperm and egg, respectively, by
mitosis. The flagellated sperm swim to the egg, and the diploid zygote divides
and grows in the archegonium and becomes the large sporophyte.
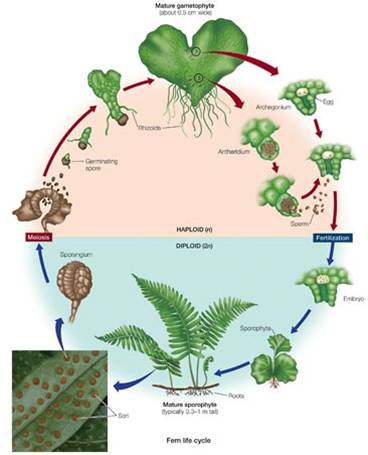
The
fern life cycle is very similar to the clubmosses; indeed, these groups were
sometimes grouped together. Ferns have a dominant sporophyte that produces true
leaves (fronds). In many species, the leaves bear sporangia underneath, which
produce spores by meiosis. The haploid spores are released, and they develope
into small, independent gametophytes that live in the leaf litter. The gametophytes
bear antheridia and archegonia, that produce sperm and egg, respectively, by
mitosis. The flagellated sperm swim to the egg, and the diploid zygote divides
and grows in the archegonium and becomes the large sporophyte.
The great swamp forests of the Carboniferous (360-286 mya, sometimes split into Mississippian and Pennsylvanian, as in the ppt) were dominated by lycopods like Lepidodendron. Tree ferns were common in the understory, and smaller lycopods and ferns were common in the herb layer, as they are today. As the large plants in these forest died, they fell into the shallow swamps and were covered by sediments. Instead of being decomposed, the organic remains were fossilized, becoming our major coal deposits and giving the period its name (Carboniferous). This huge transfer of carbon to the lithosphere changed the atmospheric balance of carbon dioxide and oxygen. The photosynthetic plants were producing lots of oxygen and were absorbing lots of carbon dioxide, coverting carbon dioxide to glucose and cellulose. Typically, the action of aerobic, decomposers would consume some of this oxygen during their respiration, and would break down the cellulose and glucose into carbon dioxide. Thus, the effects of photosynthetic organisms on the atmosphere is balanced by the effects of decomposers. However, the sequestering of this plant biomass beneath sediment meant that it wasn't metabolized and recycled; it was protected from decomposers. So, oxygen concentrations continued to rise and carbon dioxide levels dropped. Some biologists believe that the high oxygen levels may have been a factor that allowed for the largest terrestrial arthropods to survive - including a dragonfly with a 2 ft wingspan and a millipede 6 feet long. Because arthropods respire by simple diffusion, large arthropods could only receive enough oxygen to maintain their metabolic demands if the concentration gradient was so large that oxygen poured deep into tissues.
In the Permian Period, the continents united into the supercontinent of Pangaea. This land mass had a lower ratio of coastine to land area than multiple independent continents, and so most of the land mass of Pangaea was far from coastal areas. As a consequence, the terrestrial environment went through a drying period; a period which favored new groups of organisms that had evolved characteristics that would give them a selective advantage in dry environments. These organisms were the gymnosperms and the reptiles, and their adaptations were seeds and desiccation-resistant shelled eggs, respectively.
 ii.
Gymnosperms: The
gymnosperms evolved during the Carboniferous but came to dominate terrestrial
landscapes during the Permian Period, when the unification of Pangaea created
a drier terrestrial climate. They would continue to dominate through the first
part of the Mesozoic. Modern gymnosperms include the conifers (pines, fir, spruce,
hemlock, yew, redwoods,cedars, cypress, larch, etc.), the cycads, and the Ginkgo
(one relict species). Another group, the Gnetales, contains some very unusual
gymnosperms that have some characteristics of angiosperms. The gymnosperms probably
evolved from an earlier group of plants called the "Seed ferns". Seeds
ferns were the first plants to produce seeds and pollen; characteristics which
would allow their descendants to dominate terrestrial habitats. The seed ferns
are an extinct group, so we will use the modern Gymnosperms to describe the
importance of these key evolutionary innovations.
ii.
Gymnosperms: The
gymnosperms evolved during the Carboniferous but came to dominate terrestrial
landscapes during the Permian Period, when the unification of Pangaea created
a drier terrestrial climate. They would continue to dominate through the first
part of the Mesozoic. Modern gymnosperms include the conifers (pines, fir, spruce,
hemlock, yew, redwoods,cedars, cypress, larch, etc.), the cycads, and the Ginkgo
(one relict species). Another group, the Gnetales, contains some very unusual
gymnosperms that have some characteristics of angiosperms. The gymnosperms probably
evolved from an earlier group of plants called the "Seed ferns". Seeds
ferns were the first plants to produce seeds and pollen; characteristics which
would allow their descendants to dominate terrestrial habitats. The seed ferns
are an extinct group, so we will use the modern Gymnosperms to describe the
importance of these key evolutionary innovations.
- Characteristics
The seed: The plants that we have examined so far produce the diploid zygote on the female gametophyte - in the vase-like archegonium. As such, the dispersal phase of the life cycle is the spore. Spores are single celled, and although they may be dessication resistant, once they germinate and begin to grow, they must be able to immediately meet their biological needs with material they absorb from the environment. In the "seed plants" (seed ferns, gymnosperms, and angiosperms), the female gametophyte is retained on the sporophyte plant, and becomes the dispersal vessel for its offspring, the next generation of embryonic sporophyte. Here's what happens: The sporophyte produces two types of spores, large and small. Inside the ovule, a 2n cell (megasporocyte) divides by meiosis. The cytoplasm is divided unequally, producing three reduced non-functional cells and one large functional spore - the megaspore. This megaspore, nested within the ovule, divides mitotically and developes into the female gametophyte. The female gametophyte produces the egg, located at one end of the female gametophyte, within the ovule. Fertilization of the egg by sperm forms the diploid zygote. This divides to become the diploid embryo, all still surrounded by haploid gametophyte tissue, within original 2n ovule tissue. This is the seed: 2n ovule tissue (seed coat), surrounding 1n female gametophyte tissue, surrounding the 2n embryo. The gametophyte tissue stores energy in the form of fats and lipids. When the seed is dispersed, the developing embryo now has a source of nutrients to use to produce its initial roots and leaves and become established in the environment. The seed represents a way that the plant can invest MORE ENERGY in each offspring, increasing the probability of survival.
Pollen: Correlated with the production of seeds is the production of pollen. As mentioned above, the sporophyte produces two types of spores, large and small. Each small spore divides and becomes a multicellular haploid male gametophyte - a pollen grain. These pollen grains are released from the sporophyte plant, and are carried by air currents to female cones which house the female gametophyte. The pollen grain then produces sperm. Cycads and ginkgo produce flagellated sperm, but conifers and gnetales produce non-motil sperm. In either case, the sperm is already at the egg. Obviously, the evolution of pollen, that carries sperm through the environment, meant that water was no longer required as a vehicle for reproduction. Plants were now free to colonize uplant areas away from water.
- Diversity:
There are four groups of living gymnosperms. The cycads are the most primitive group, perhaps reminiscent of the seed ferns from which the gymnosperms evolved. They still have flagellated sperm (an ancestral trait), and stiff, tough, frond-like leaves. Some Cycads even bear megaspores on leaves like the ferns do, although the male microspores and pollen are produced in cones. They also have an unbranched stem that is similar to tree ferns. They do produce true seeds, however, and thus are true (though primitive) gymnosperms. The Ginkgo tree is an unusual species - it is the only species in its entire Order (Ginkgoales) - the last species remaining from a previously diverse group. It also has the primitive trait of flagellated sperm. Pollen is produced in cones, but megaspores are produced in fruit-like structures. The third group - Gnetales, - is the sister group to conifers and is a diverse collection of odd plants. The most diverse group of gymnosperms on the planet today is the conifers. This group includes ~600 of the roughly 900 species of gymnosperms (compared to 300,000 species of flowering plants!!). The group includes the largest plants on earth - the tall redwoods and the massive sequioas. It also includes the oldest plants on the planet - some redwoods and bristlecone pines are over 5000 years old. Conifers include pines, fir trees, spruce, larch, yews, cypress, redwoods and sequioas, hemlock, cedar, and juniper. Conifers, like other gymnosperm groups, arose in the Carboniferous and radiated in the Permian when the climate became drier. Conifers, in particular, radiated during the Mesozoic, filling plant niches emptied by the great Permian extinction. Conifers continue to dominate dry environments, today. They dominate dry coastal habitats where sandy soils reduce water availability, they dominate dry mountain slopes, and they dominate cold habitats at high elevations and high latitudes where water is often frozen and unavailable. Indeed, the boreal forest - one of the largest terrestrial biomes on the planet, is overwhelmingly dominated by conifers. So, although their diversity has diminished and although they have been numerically dominated by flowering plants, gymnosperms continue to be very important components of many terrestrial habitats.
 |
 |
 |
 |
| Cycads have large, fern-like leaves | Ginkgo is a single 'living fossil' species | Gnetales are just weird | Conifers still dominate many habitats |
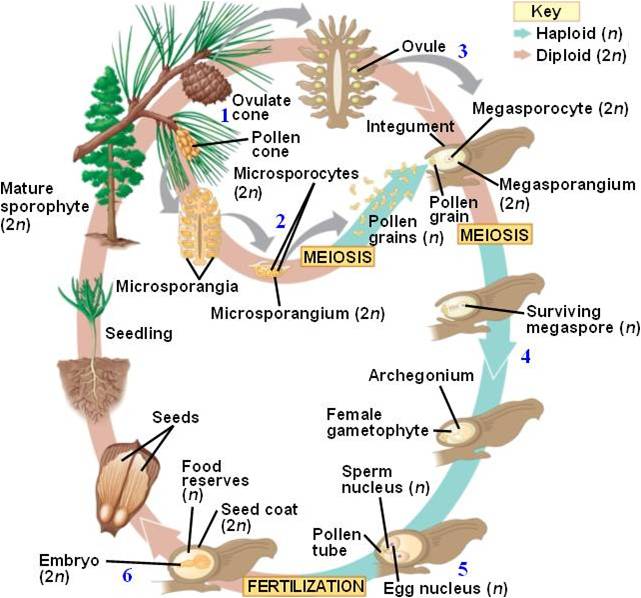 Life
cycle: We will use a pine tree (conifer) as our example of the gymnosperm
life cycle. Consider a mature pine tree - it is the diploid sporophyte generation.
It produces two types of cones, male and female. In the male cones, in compartments
on each "scale", 2n microsporocyte cells go through meiosis and produce
4 equally sized haploid microspores. Each microspore divides by mitosis and
develops into a mature haploid male gametophyte - a pollen grain. Each pollen
grain then produces haploid non-motil sperm by mitosis. Pine trees also have
larger, woody, female cones. In ovules on each scale of these cones, meiosis
occurs and produces a single large haploid spore - the megaspore. The megaspore
divides by mitosis, producing the multicellular haploid female gametophyte.
This megagametophyte produces the egg. There is a pore at one end of the ovule
called the micropyle. When the gametophyte is sexually mature, it secretes a
drop of fluid called the "pollen droplet". Pollen is released from
the male cones and wafts into the female cones. It gets stuck in the fluid droplet,
and as the droplet dries the pollen grain is pulled into the ovule. Inside the
ovule, a tube cell bores through the pollen grain, followed by sperm cells.
Fertilization takes place, forming a diploid zygote within the haploid female
gametophyte tissue. So, there is a diploid embryo, surrounded by haploid gametophyte
tissue, surrounded by a thin covering (seed coat) of grand-parental sporophyte
tissue. This three-layered structure is the seed, and it can be dispersed to
a new environment and gain a footing in that habitat by using the energy packet
along in the female gametophyte tissue.
Life
cycle: We will use a pine tree (conifer) as our example of the gymnosperm
life cycle. Consider a mature pine tree - it is the diploid sporophyte generation.
It produces two types of cones, male and female. In the male cones, in compartments
on each "scale", 2n microsporocyte cells go through meiosis and produce
4 equally sized haploid microspores. Each microspore divides by mitosis and
develops into a mature haploid male gametophyte - a pollen grain. Each pollen
grain then produces haploid non-motil sperm by mitosis. Pine trees also have
larger, woody, female cones. In ovules on each scale of these cones, meiosis
occurs and produces a single large haploid spore - the megaspore. The megaspore
divides by mitosis, producing the multicellular haploid female gametophyte.
This megagametophyte produces the egg. There is a pore at one end of the ovule
called the micropyle. When the gametophyte is sexually mature, it secretes a
drop of fluid called the "pollen droplet". Pollen is released from
the male cones and wafts into the female cones. It gets stuck in the fluid droplet,
and as the droplet dries the pollen grain is pulled into the ovule. Inside the
ovule, a tube cell bores through the pollen grain, followed by sperm cells.
Fertilization takes place, forming a diploid zygote within the haploid female
gametophyte tissue. So, there is a diploid embryo, surrounded by haploid gametophyte
tissue, surrounded by a thin covering (seed coat) of grand-parental sporophyte
tissue. This three-layered structure is the seed, and it can be dispersed to
a new environment and gain a footing in that habitat by using the energy packet
along in the female gametophyte tissue.
With seed fern and conifer evolution, plants had finally become independent of an aquatic environment. No life stage needed liquid water as an environment, and these plant species could colonize the drier areas that were increasingly abundant as the Permian and Mesozoic progressed.
iii. Angiosperms: The angiosperms are the flowering plants; they are the most diverse group of plants on the planet today, represented by approximately 300,000 species and comprising 90% of all plant species. Angiosperms evolved early in the mesozoic, but radiated in the early Cretaceous Period - the last period of the Mesozoic Era. Although their diversity declined as a consequence of the meteor impact 65 mya (known as the K-T boundary because it is used to separate the Cretaceous ("K" - yeah, I know... but Cambrian got "C") of the Mesozoic from the Tertiary ("T") of the Cenozoic Era), they rebounded and dominated most terrestrial habitats. Like their sister group the gymnosperms, they have seeds and pollen. However, they also have two additional distinguishing derived characteristics - flowers and fruits.
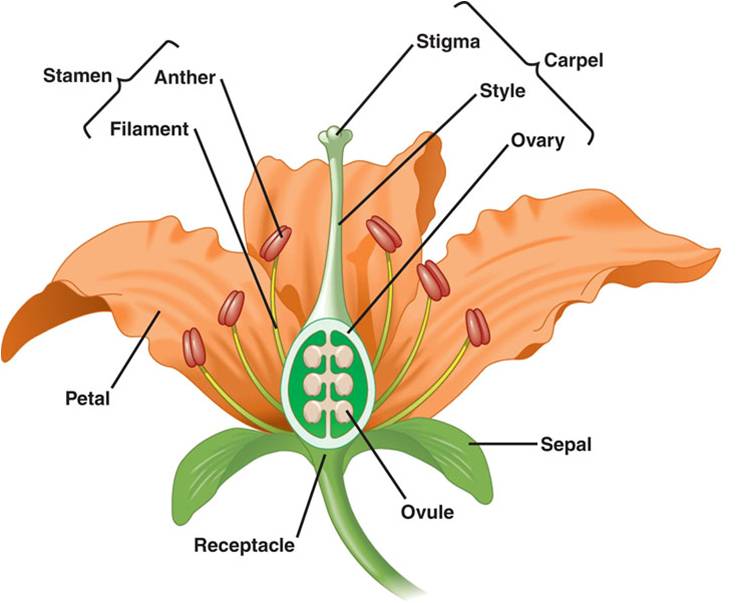 Flowers:
A flower is a stem-like structure that bears whorls of leaf-like appendages.
The lowest whorl on this reproductive "twig" is the calyx, composed
of leaf-like sepals that form a protective shealth surrounding the flower bud.
The second whorl of leaf-like appendages is the corolla, containing the leaf-like
petals. The third whorl contains the stamens. Each stamen has a filament that
bears an anther at the end. The anther is where meiosis occurs and haploid microspores
develop into pollen grains. At the center of the flower is the pistil, or carpel.
This is the female portion of the flower, consisting of the stigma (a sticky
surface that receives pollen), the style (a stalk connecting the stigma to the
ovary) and the ovary. The ovary has compartments that contain the ovules. The
structure of flowers can vary tremendously, and may lack carpels, petals, pistils,
or stamens. The number of the parts may vary, also, and 'composite' flowers
may contain thousands of flowers growing together as one big inflorescence -
with some flowers only bearing petals, and other flowers only bearing stamens
or ovaries. A daisy is a good example of a composite - the petals are actually
the product of single 'ray' flowers that each ONLY bear one petal. The yellow
center of a daisy is composed of hundreds of individual 'disk' flowers that
don't have petals but do have stamens and pistils.
Flowers:
A flower is a stem-like structure that bears whorls of leaf-like appendages.
The lowest whorl on this reproductive "twig" is the calyx, composed
of leaf-like sepals that form a protective shealth surrounding the flower bud.
The second whorl of leaf-like appendages is the corolla, containing the leaf-like
petals. The third whorl contains the stamens. Each stamen has a filament that
bears an anther at the end. The anther is where meiosis occurs and haploid microspores
develop into pollen grains. At the center of the flower is the pistil, or carpel.
This is the female portion of the flower, consisting of the stigma (a sticky
surface that receives pollen), the style (a stalk connecting the stigma to the
ovary) and the ovary. The ovary has compartments that contain the ovules. The
structure of flowers can vary tremendously, and may lack carpels, petals, pistils,
or stamens. The number of the parts may vary, also, and 'composite' flowers
may contain thousands of flowers growing together as one big inflorescence -
with some flowers only bearing petals, and other flowers only bearing stamens
or ovaries. A daisy is a good example of a composite - the petals are actually
the product of single 'ray' flowers that each ONLY bear one petal. The yellow
center of a daisy is composed of hundreds of individual 'disk' flowers that
don't have petals but do have stamens and pistils.
Flowers evolved to exploit animals for pollen transfer. Wind is capricious - the direction changes, the intensity changes, and even on a windy day only a small fraction of the pollen produced in a male cone will be lucky enough, just by chance, to travel to a female cone. For this reason, gymnosperms typically produce alot of pollen. By contrast, animals move through the environment non-randomly in search of food, shelter, and mates; and all animals can learn. So, by associating pollen with attractants (nectar, fragrance, and colorful floral displays) plants teach animals to visit flowers and increase the frequency of effective pollen transfer. Animals visiting an attractive flower are rewarded with food and quickly learn the association; they then travel in "bee-lines" from one flower to another seeking food, transfering pollen non-randomly and efficiently. This would be particularly useful for plants in the understory of a forest, where wind is minimal. The first angiosperms may have been shrubby, understory species. Animal pollination would also be very adaptive in tropical rainforests or deserts, where members of a single plant species may be very far away (decreasing the chances that a random procees like wind pollination would effectively find a 'mate').
As some angiosperms evolved into taller forms where wind could be used effectively, some were selected to use wind as a pollen-dispersal mechanism - with correlated reductions in flower size. So, wind-pollinated flowering trees (oaks, maples, etc.) and wind-pollinated field plants (open fields are windy environments, too) like grasses often have very small flowers with no colorful petals or sepals. Because they are not attracting animals for pollen dispersal, selection favors organisms that do not invest alot of energy in these structures.
 Plants
can increase the efficiency of pollen transfer even further by limiting flower
visitation to one type of pollinator. A specialized pollinator that visits just
one type of flower is more likely to transfer pollen between plants of the same
species. Likewise, flowers visited by just one type of pollinator are likely
to have nectar present when that pollintor visits. So, "pollination syndromes"
have evolved, representing flower traits that are associated with a particular
type of pollinator. For example, many flowers are red and tubelike. These flowers
are often pollinated by hummingbirds, which see into the red end of the spectrum
better than most insects and have a long beak that can access the nectar at
the base of the flower. Butterfly flowers are often composites (like daisies,
and zinnias), or are clustered in groups (butterfly bush). This allows the butterfly
to land on a cluster and probe many individual flowers at a time with their
long proboscis. Moth and bat pollinated flowers open at night. They are usually
white and very fragrant, attracting their pollinators with scent. Some plants
like skunk cabbage produce flowers that smell like rotting flesh. This attracts
flies that act as pollinators. Some of the most elaborate contrivances by which
plants limit pollination to a single species of pollinator are seen in the orchid
family. Some orchids produce a scent that mimics the sexual pheromones of females
of specific species of solitary wasps. When males perceive the scent, they fly
up the concentration gradient to the flower. In some species, the flower also
mimics the shape and coloration of the female wasp! The deception is so good
that the male attempts to mate with the flower. It gets dusted with pollen,
and flies off. While this may be frustrating for the wasp (who knows), it can't
NOT fly down the next pheromone trail it encounters or it won't mate. So, if
it flies down another trail and is tricked by another plant, pollen is transferred.
The bucket orchid is another strange plant. Two petals have fused into a bucket
that collect water. Pollinators fall in the bucket, and the only exit is through
a tight squeezeway where they are dusted with pollen.
Plants
can increase the efficiency of pollen transfer even further by limiting flower
visitation to one type of pollinator. A specialized pollinator that visits just
one type of flower is more likely to transfer pollen between plants of the same
species. Likewise, flowers visited by just one type of pollinator are likely
to have nectar present when that pollintor visits. So, "pollination syndromes"
have evolved, representing flower traits that are associated with a particular
type of pollinator. For example, many flowers are red and tubelike. These flowers
are often pollinated by hummingbirds, which see into the red end of the spectrum
better than most insects and have a long beak that can access the nectar at
the base of the flower. Butterfly flowers are often composites (like daisies,
and zinnias), or are clustered in groups (butterfly bush). This allows the butterfly
to land on a cluster and probe many individual flowers at a time with their
long proboscis. Moth and bat pollinated flowers open at night. They are usually
white and very fragrant, attracting their pollinators with scent. Some plants
like skunk cabbage produce flowers that smell like rotting flesh. This attracts
flies that act as pollinators. Some of the most elaborate contrivances by which
plants limit pollination to a single species of pollinator are seen in the orchid
family. Some orchids produce a scent that mimics the sexual pheromones of females
of specific species of solitary wasps. When males perceive the scent, they fly
up the concentration gradient to the flower. In some species, the flower also
mimics the shape and coloration of the female wasp! The deception is so good
that the male attempts to mate with the flower. It gets dusted with pollen,
and flies off. While this may be frustrating for the wasp (who knows), it can't
NOT fly down the next pheromone trail it encounters or it won't mate. So, if
it flies down another trail and is tricked by another plant, pollen is transferred.
The bucket orchid is another strange plant. Two petals have fused into a bucket
that collect water. Pollinators fall in the bucket, and the only exit is through
a tight squeezeway where they are dusted with pollen.
Fruits:Many angiosperms also evolved mechanisms to use animals to disperse their seeds. The ancestral condition within the group is probably the production of fleshy, ovary tissue that surrounds the carpels. Like nectar, this fleshy fruit tissue serves as a bribe, encouraging animals to eat the fruits and thereby disperse the seeds. And since animals tend to show a preference for a given habitat type, the seeds get transferred to another place with similar environmental conditions (rather than being strewn haphazardly across the environment by wind). Other fruits are dry - like acorns, grains, burrs, and some beans. If these are small, like in grasses, or have wings like maple samaras, or have feathery structures like milkweed, then the fruit can be dispersed by the wind. Some, like cocklebur, burdock, and goatshead, evolved hooks on their seed pods that could tangle in mammal fur. Some fruits are pea-pod like, and when the pod dries, the seam bursts open and the seeds are "shot out". This is called 'ballistic' dispersal. So, regardless of whether the fruit is animal or wind dispersed, plants have evolve many different ways to get their seeds dispersed and away from the competitive environment beneath the parent plant.
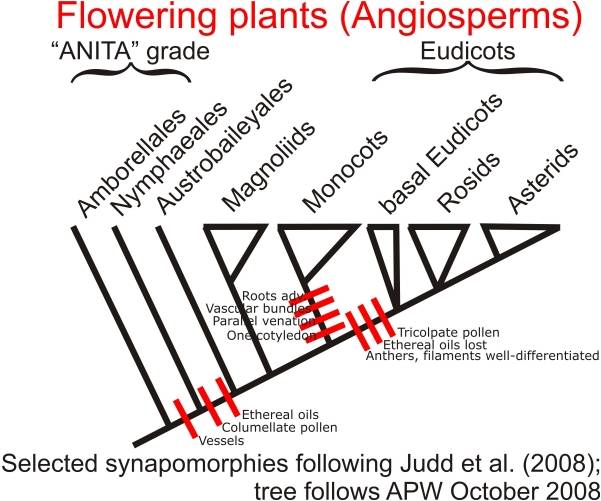 Diversity:
The 300,000 angiosperm species have radiated into such a wide variety of shapes
and behaviors that the evolutionary relationships among major groups (which
each contain lots of diversity) is often difficult. However, several groups
do fall out, especially with molecular analyses. Two ancient, basal groups are
the Amborellales ( a small group of a few very primitive species) and the Nymphaeales
(water lilies and their kin). Another ancient group is the Magnoliales, including
the Magnolias and the laurels. Two other large and diverse groups are the monocots
(including grasses, palms, lilies, and orchids), and the dicots (including 70%
of all plants, from roses to oaks, from buttercups to maples). Monocots have
one embryonic leaf (cotyledon), while dicots have two. Also, monocots have leaves
with parallel veins, while dicots usually have leaves with a network of radiating
veins. Monocots typically have flower parts in multiples of 3's, while dicots
have flower parts in multiples of 4's or 5's.
Diversity:
The 300,000 angiosperm species have radiated into such a wide variety of shapes
and behaviors that the evolutionary relationships among major groups (which
each contain lots of diversity) is often difficult. However, several groups
do fall out, especially with molecular analyses. Two ancient, basal groups are
the Amborellales ( a small group of a few very primitive species) and the Nymphaeales
(water lilies and their kin). Another ancient group is the Magnoliales, including
the Magnolias and the laurels. Two other large and diverse groups are the monocots
(including grasses, palms, lilies, and orchids), and the dicots (including 70%
of all plants, from roses to oaks, from buttercups to maples). Monocots have
one embryonic leaf (cotyledon), while dicots have two. Also, monocots have leaves
with parallel veins, while dicots usually have leaves with a network of radiating
veins. Monocots typically have flower parts in multiples of 3's, while dicots
have flower parts in multiples of 4's or 5's.
Life Cycle: The generalized life cycle of angiosperms is similar to gymnosperms. The diploid sporophyte produces two types of spores. In angiosperms, however, these can be born by the same structure (perfect hermaphroditic flowers), or different flowers on the same plant (monoecious) or on different plants (dioecious). In the anther, 2n microsporocytes divide by meiosis and produce 4 microspores. Each microspore divides by mitoisis and developes into a pollen grain. Pollen is released from the anther - often on to the body of an animal pollinator visiting the flower. In the ovule, a diploid megasporocyte divides unequally, producing 3 nonfunctional cells and one functional haploid megasporocyte. The nucleus in this cell divides by mitosis through three cycles, producing 8 nuclei. Three aggregate near each pole, and two agregate in the middle of the ovule. The central cell at one end becomes the egg.
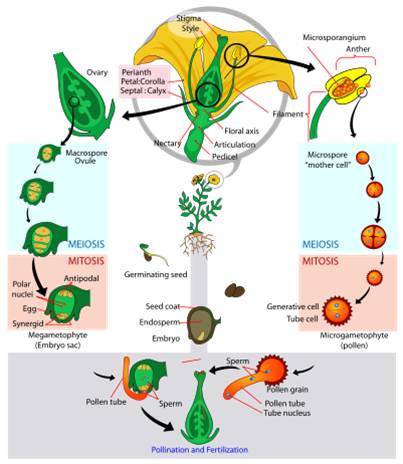 Pollen
lands on the stigma, and one tube cell secretes enzymes that dig a "pollen
tube" through the style to the ovule. It is followed down the tube by two
sperm cells. When they reach the opening of the ovule, one sperm cell fertilizes
the egg and forms the diploid zygote. The other sperm cell fuses with the TWO
polar nuclei in the middle of the ovule, forming a triploid nucleus. This "double
fertilization" is unique to the angiosperms. The triploid nucleus divides,
forming triploid 'endosperm' tissue within the ovule that surrounds the developing
diploid enbryo and provides nutrition. So again, the seed contains three layers
- the ovule of the original diploid sporophyte, the triploid endosperm, and
the diploid embryo.
Pollen
lands on the stigma, and one tube cell secretes enzymes that dig a "pollen
tube" through the style to the ovule. It is followed down the tube by two
sperm cells. When they reach the opening of the ovule, one sperm cell fertilizes
the egg and forms the diploid zygote. The other sperm cell fuses with the TWO
polar nuclei in the middle of the ovule, forming a triploid nucleus. This "double
fertilization" is unique to the angiosperms. The triploid nucleus divides,
forming triploid 'endosperm' tissue within the ovule that surrounds the developing
diploid enbryo and provides nutrition. So again, the seed contains three layers
- the ovule of the original diploid sporophyte, the triploid endosperm, and
the diploid embryo.
6. Summary:
Over the course of plant evolution, organisms evolve characteristics that provided a selective advantage in progessively more desiccating environments, allowing plants to colonize new habitats unoccupied by other plants. The non-tracehophytes evolved a cuticle, and could colonize land and harvest more sunlight than their algal ancestors. Club mosses evolved vascular tissue and a dominant sporophyte, allowing them to grow tall and gain a competitive advantage over smaller non-tracheophytes. Ferns evolved more complex leaves, improving the transport of water and sugars into and out of leaves. The evolution of seeds, the reduction of the the gametophyte, and the elimination of swimming sperm allowed seed ferns and their descendants - the gymnosperms and the angiosperms - to colonize upland areas away from water. Conifers still dominate in the boreal forests of high latitudes; however, the angiosperms dominate in almost all other terrestrial habitats. Their innovations of flowers and fruits exploit animals in the environment, and improve the efficiency of pollen transport and seed dispersal. The evolution of plants is an interesting story of adapting to life on land, to take advantage of the abundant sunlight, oxygen, and mineral nutrients found in this habitat. The are an extraordinarily important, interesting, diverse, and beautiful group of living organisms.
1. Overview:
The other major opisthokont group is the Animalia. Animals are distinguished as multicellular, heterotrophic eukaryotes that lack a cell wall. Obviously, the lack of a cell wall distinguishes them from the other multicelluar, heterotropic eukaryotes - the fungi. There is genetic and morphological evidence that animals evolved from choanoflagellate ancestors - probably between 900-700 mya. The oldest animal fossils unearthed to date were found in Australia, in August 2010. They are thought to be sponges, and they date to 650 mya. Sponges are the most 'primitive' group of animals, with the most direct ties to the choanoflagellates, so their early appearance in the fossil record corroborates phylogenetic hypotheses.
 Animal
fossils become much more abundant in the fossil record 550 mya. Although there
are older fossils of soft -bodied jellyfish and worms, the fossil record seems
to 'explode' about 550 mya - an observation that led early Paleotologists to
demarcate this as the beginning of the Paleozoic Era. This explosion probably
occurred because some animals - notably arthropods and molluscs - evolved a
protective body covering (exoskeleton or shell, respectively) that protected
them against the stinging cnidarian predators. The radiation of these groups
left a disproportionate mark on the fossil record because hard parts fossilize
much more readily than soft tissue. So, in part, the 'Cambrian Explosion' is
probably due to sampling bias - organisms evolved that were more likely to fossilize,
so we see more fossils. The evolution of arthropods in the Cambrian led to an
explosion of diversity that is still occurring today - over 85% of the animal
species named to date are Arthropods. In the figure to the right, we see an
artists depiction of the Cambrian Period, with a variety of sponges, cnidarians,
molluscs, and arthropods, including the large predatory arthropod, Anomalocaris
- that grasped other animals with its raptorial claws and crushed them in their
circular jaws.
Animal
fossils become much more abundant in the fossil record 550 mya. Although there
are older fossils of soft -bodied jellyfish and worms, the fossil record seems
to 'explode' about 550 mya - an observation that led early Paleotologists to
demarcate this as the beginning of the Paleozoic Era. This explosion probably
occurred because some animals - notably arthropods and molluscs - evolved a
protective body covering (exoskeleton or shell, respectively) that protected
them against the stinging cnidarian predators. The radiation of these groups
left a disproportionate mark on the fossil record because hard parts fossilize
much more readily than soft tissue. So, in part, the 'Cambrian Explosion' is
probably due to sampling bias - organisms evolved that were more likely to fossilize,
so we see more fossils. The evolution of arthropods in the Cambrian led to an
explosion of diversity that is still occurring today - over 85% of the animal
species named to date are Arthropods. In the figure to the right, we see an
artists depiction of the Cambrian Period, with a variety of sponges, cnidarians,
molluscs, and arthropods, including the large predatory arthropod, Anomalocaris
- that grasped other animals with its raptorial claws and crushed them in their
circular jaws.
Animals are a fairly recent addition to the living world. They are not very genetically or metabolically diverse when compared to prokaryotes or protists that have much longer evolutionary histories. However, animals are structurally diverse. They have radiated into a wide variety of shapes through changes in developmental patterning. These different shapes have allowed them to exploit a wide range of habitats and resources. Animals are classified into phyla based on their shape or body plan. As heterotrophs, these differences often correlate with differences in feeding structures or locomotory structures that they use to capture food.
From our vantage point as large terrestrial organisms, we have a very skewed view of animal diversity. This is represented by our casual division of animals into 'vertebrates' (like us) and 'invertebrates' (other stuff not like us). These are not phylogenetic groups. The Vertebrata is one sub-phylum of the phylum Chordata, which is one of 32 phyla of animals. The two other sub-phyla of Chordates, and all the other 31 phyla, are 'invertebrate' - and they obviously represent the bulk of animal diversity. Indeed, 95% of all animal species are 'invertebrates', with 85%, as mentioned, occurring in the single phylum Arthropoda. In addition, most animal phyla are exclusively marine - so we rarely encounter them. As you look through the groups pictured at the top of the page (the protostome phyla), you'll see that most animals are rather simple, bilaterally symmetrical, worm-like organisms with different feeding structures.
We are only going to consider the major phyla of animals here, those representing important evolutionary innovations in the group, or representing important radiations.
2. Development
 One
of the defining features in the animals is their method of early embryonic development.
The subsequent differences that develop in shape occur after these initial stages
take place. The union of sperm and egg form a diploid zygote. the zygote divides
by mitosis, forming a solid ball of cells called the morula. The cells in the
morula migrate, moving out from under other cells until they form a hollow ball
of cells called the blastula. Then, one side of the blastula migrates in towards
the center of the embryo. This process of infolding, or invagination, creates
the gastrula. The gastrula has two layers of cells: one layer is on the outside
(ectoderm) and the other (endoderm) is on the inside, lining the newly-formed
cavity the infolding produced. This cavity becomes the gut in all animals. The
hole that connects the gut with the environment becomes either the mouth or
anus of the mature animal. Although sponge development is slight different,
they still form these typical stages. In all other animals, from cnidarians
to cats, the process is fundamentally the same. Differences in shape emerge
as the embryo developes from this gastrula.
One
of the defining features in the animals is their method of early embryonic development.
The subsequent differences that develop in shape occur after these initial stages
take place. The union of sperm and egg form a diploid zygote. the zygote divides
by mitosis, forming a solid ball of cells called the morula. The cells in the
morula migrate, moving out from under other cells until they form a hollow ball
of cells called the blastula. Then, one side of the blastula migrates in towards
the center of the embryo. This process of infolding, or invagination, creates
the gastrula. The gastrula has two layers of cells: one layer is on the outside
(ectoderm) and the other (endoderm) is on the inside, lining the newly-formed
cavity the infolding produced. This cavity becomes the gut in all animals. The
hole that connects the gut with the environment becomes either the mouth or
anus of the mature animal. Although sponge development is slight different,
they still form these typical stages. In all other animals, from cnidarians
to cats, the process is fundamentally the same. Differences in shape emerge
as the embryo developes from this gastrula.
3. Body Plans
 Animals
exibit a wide range of shapes, but these shapes can be classified in three major
groups. First, the sponges are asymmetrical. Second, the cnidarians and ctenophores
are 'radially symmetrical. This means that, if you are looking down on the organism
from the top, you could cut the organism is half along any axis passing through
the midpoint and the two halves would be be the same. The rest of the animals
are bilaterally symmetrical. This means that there is only one plane of division
that will create to symmetrical 'sides' (bi-lateral = two sides). Given the
fact that the vast majority of animal phyla are bilaterally symmetrical, we
can hypothesize that this evolutionary innovation was functionally important
- allowing animals to work "better", somehow. But how?
Animals
exibit a wide range of shapes, but these shapes can be classified in three major
groups. First, the sponges are asymmetrical. Second, the cnidarians and ctenophores
are 'radially symmetrical. This means that, if you are looking down on the organism
from the top, you could cut the organism is half along any axis passing through
the midpoint and the two halves would be be the same. The rest of the animals
are bilaterally symmetrical. This means that there is only one plane of division
that will create to symmetrical 'sides' (bi-lateral = two sides). Given the
fact that the vast majority of animal phyla are bilaterally symmetrical, we
can hypothesize that this evolutionary innovation was functionally important
- allowing animals to work "better", somehow. But how?
Bilateral symmetry does more than create two sides; it also means that there is a top and bottom, and a front and back. As heterotrophs, most animals have to moving through the environment in search for their food. Having a 'front' would favor the concentration of sensory systems at this end - so that the animal could perceive the environment that it was about to enter and search for food and avoid predators. So, bilateral symmetry is associated with the formation of a head and the evolution of a brain for processing all this sensory information and directing locomotion. It is no coincidence, therefore, that bilateral symmetry allowed some animals to "get a-head". hahaha.
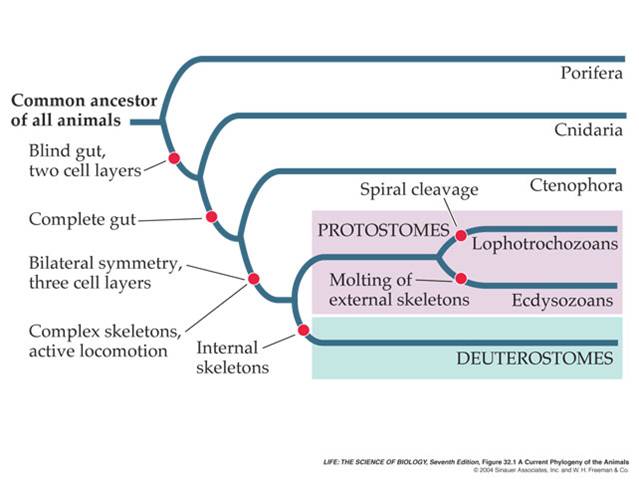 4.
Phylogeny
4.
Phylogeny
These developmental patterns are reflected by animal phylogeny. Sponges are the simplest animals; they retain the collar cells (choanocytes) that are so similar to the choanoflagellates, and the cells in sponges do not form true tissue layers (where the cells are bound together in a matrix of secreted proteins). The remaining animals are distinguished by having only two true tissues (ecto and endoderm) or three (with mesoderm). In addition, the more primitive animals are radially symmetrical, while the animals with three tissue layers are bilaterally symmetrical. These bilateral animals are grouped by whether the blastopore forms the mouth ('protostomes') or anus ('deuterostomes'). The protostomes are further classified by how their larvae develop or how they grow: lophotrochozoans have particualr larval stages and grow by adding segments in some cases, whereas the ecdysozoans have a cuticle/exoskeleton and must shed this in order to grow. We will review these patterns as we move through the major animal phyla.
5. Major Phyla
a. Phylum Porifera: Sponges
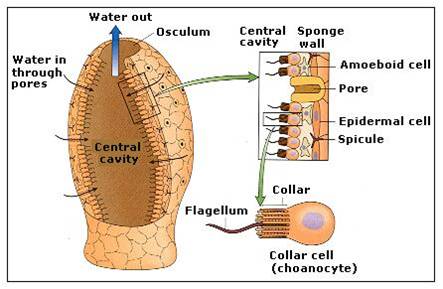 Sponges
consist of specialized but loosely integrated cells; the cells are not
anchored in a layer of protein (known as a basement membrane) as in
other animals. So, curiously, some sponges can be 'disintegrated' in a blender
and will then re-assemble into many small sponges, with cell types reintegrating
into their appropriate relationship. These aggregates are asymmetrical, but
the fundamental filtering unit is often vase-like. The cells in a vase unit
form two layers; the inside layer consists of choanocytes that are very similar
to free-living choanoflagellates. The beating of the flagella create a current
that pulls water through pore cells into the central chamber of the sponge -
water exits through the top hole or 'osculum'. The choanocytes feed by ingesting
organic material by phagocytosis. The outer cell layer is the 'epidermis'. The
epidermis receives nutrients from the choanocytes in an interesting way: the
food that is absorbed by the inner layer of choanocytes is passed to amoebocytes
- motile cells that live in the space between the cell layers. This space, the
mesohyl, contains flexible spongin proteins, and may contain rigid spicules
of silca. The ameobocyte crawls back and forth, transferring food from the chaoanocytes
to the epidermal cells. Sponges are filter feeders. They reproduce asexually
by fragmentation (pieces can break off and settle elsewhere), and by sexual
reproduction.
Sponges
consist of specialized but loosely integrated cells; the cells are not
anchored in a layer of protein (known as a basement membrane) as in
other animals. So, curiously, some sponges can be 'disintegrated' in a blender
and will then re-assemble into many small sponges, with cell types reintegrating
into their appropriate relationship. These aggregates are asymmetrical, but
the fundamental filtering unit is often vase-like. The cells in a vase unit
form two layers; the inside layer consists of choanocytes that are very similar
to free-living choanoflagellates. The beating of the flagella create a current
that pulls water through pore cells into the central chamber of the sponge -
water exits through the top hole or 'osculum'. The choanocytes feed by ingesting
organic material by phagocytosis. The outer cell layer is the 'epidermis'. The
epidermis receives nutrients from the choanocytes in an interesting way: the
food that is absorbed by the inner layer of choanocytes is passed to amoebocytes
- motile cells that live in the space between the cell layers. This space, the
mesohyl, contains flexible spongin proteins, and may contain rigid spicules
of silca. The ameobocyte crawls back and forth, transferring food from the chaoanocytes
to the epidermal cells. Sponges are filter feeders. They reproduce asexually
by fragmentation (pieces can break off and settle elsewhere), and by sexual
reproduction.
b. Phylum Cnidaria: Hydra, anemones, corals, jellyfish
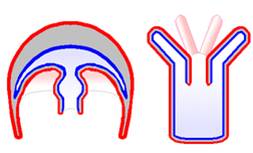 Although
the cnidarians may seem to be quite different from one another, the body plans
of jellyfish ('medusa' plan) and anemones ('polyp' plan) are actually very similar.
All cnidarians have a rather simple plan of two true tissue layers separated
by a gel/protein filled space (mesoglea). The outer layer covers the tentacles,
which have cnidocytes. These are cells that house a harpoon-like structure.
Any bump or brush and the harpoon is deployed, along with toxins that paralyze
the prey. The tentacles contract and draw the prey into the gut cavity, where
extracellular digestion occurs. Emzymes are produced by endodermal tissue and
these enzymes are secreted into the gut cavity. The products of this digestion
are absorbed by cells by phagocytosis, so there is both intra and extracellular
digestion. As in sponges, there are ameobocytes that transfer nutrients from
the endodermis to the ectodermis. The inner cell layer, the endoderm, lines
the gut cavity. In cnidarians, digestion is both extracellular and intracellular.
Cnidarians have a 'nerve net' - a decentralized organization of nerve-like cells
that coordinate the movement of tentacles. Some cnidarians are exclusively asexual,
but most can reproduce sexually.
Although
the cnidarians may seem to be quite different from one another, the body plans
of jellyfish ('medusa' plan) and anemones ('polyp' plan) are actually very similar.
All cnidarians have a rather simple plan of two true tissue layers separated
by a gel/protein filled space (mesoglea). The outer layer covers the tentacles,
which have cnidocytes. These are cells that house a harpoon-like structure.
Any bump or brush and the harpoon is deployed, along with toxins that paralyze
the prey. The tentacles contract and draw the prey into the gut cavity, where
extracellular digestion occurs. Emzymes are produced by endodermal tissue and
these enzymes are secreted into the gut cavity. The products of this digestion
are absorbed by cells by phagocytosis, so there is both intra and extracellular
digestion. As in sponges, there are ameobocytes that transfer nutrients from
the endodermis to the ectodermis. The inner cell layer, the endoderm, lines
the gut cavity. In cnidarians, digestion is both extracellular and intracellular.
Cnidarians have a 'nerve net' - a decentralized organization of nerve-like cells
that coordinate the movement of tentacles. Some cnidarians are exclusively asexual,
but most can reproduce sexually.
The "Bilateria"
The remaining animal phyla are bilaterally symmetrical organisms. Their symmetry is encoded by Hox genes that establish the anterior-posterior axis of the body. As mentioned previously, bilateral symmetry is advantageous because it establishes a polarity to the animal - and sense systems and neural integration can be concentrated at the anterior end.
The bilaterally symmetrical animals have three true tissue layers (triploblastic), adding mesoderm to endo- and ectoderm. They form two major clades, distinguished by whether the blastopore developes into the mouth (Protostomes) or the anus (Deuterostomes) in the adult. The protostomes are further divided by other developmental characteristics. One major clade, the Ecdysozoa, must shed a hard cuticle or exoskeleton in order to grow. The other clade, the Lophotrochozoans, is distinguished by having either a rake-like feeding apparatus ('lophophore' - seen in some of the pictures in the lower two rows, above), or a specific larval stage known as a 'trochophore'. As you can see from the phylogeny above, there are many groups whose phylogenic relationships are unknown at this time.
Protostomes: Lophotrochozoans
c. Phylum Platyhelminthes: Flatworms
 The
flatworms are a rather primitive group of bilaterally symmetrical organisms
that maintain some of the basic characteristics of cnidarians. Like cnidarians,
they have a digestive cavity with only one opening; they do not have a one-way
digestive tract. However, the gut of flatworms is highly bifurcated and convoluted,
with pockets that radiate out into the deep tissue on the sides of the animal.
This is probably adaptive in this organism because it insures that no cell is
too far from the gut (and digested nutrients). Flatworms also have ameobocytes
like sponges and cnidarians, too; these transport nutrients between the endodermal
cells lining the gut and the rest of the tissues in the animal. Digestion is
both intracellular and extracellular, as in cnidarians. The major evolutionary
innovations in this group are associated with bilaterality. The nervous system
and sensory organs (eyespots and chemoreceptive auricles) are concentrated towards
the anterior end of the animal, creating a head (of sorts). Flatworms are a
diverse group, with free-living and parasitic forms. Planaria live in marine,
freshwater, and terrestrial environments. Flukes are parasitic, like the liver
flukes and blood flukes (Schistosomes) we saw in lab. Most of these species
have at least one intermediate host and a complex life cycle. At some point
in the life cycle there is usually an asexual stage in which thousands of offspring
can be produced asexually. This is particularly important for parasites that
infect different hosts in sequence, because the probability of finding new hosts
is low. Tapeworms are another group of parastic flatworms. Over 1000 species
have been described, and every vertebrate species is infected by at least one
species of tapeworm. Humans are infected by three, with sheep, pigs, cattle,
and fish acting as intermediate hosts. Tapeworms are weird. Humans ingest living
cysts in undercooked beef, lamb, pork, and fish. The immature worm hatches from
the cysts and attached to the wall of the intestine with the hooks on the scolex
- the head segment. As the worm matures and grows, it produces body segments
called proglottids. Each proglottid contains male and female reproductive organs.
As the worm creates new proglottids behind the head, older proglottids that
are farther from the head mature. The worm can be over 12m long in humans; some
grow to over 30m in length! Each proglottid is metabolically independent, absorbing
nutrients by diffusion, directly from the hosts intestinal tract. The high SA/V
ratio of these flatworms aids in this parasitic lifestyle. Each proglottid produces
egg and sperm; sperm are received by proglottids and the development of fertilized
eggs occurs in each proglottid. Eventually, the mature proglottids are shed
by the worm, and are shed from the host in feces. The proglottids are eaten
by the secondary host (sheep, pigs, cattle, and fish). The larvae hatch, bore
through the intestinal wall, and migrate to muscle tissue where they encyst.
Consumption of undercooked meat by another person completes the life cycle.
The
flatworms are a rather primitive group of bilaterally symmetrical organisms
that maintain some of the basic characteristics of cnidarians. Like cnidarians,
they have a digestive cavity with only one opening; they do not have a one-way
digestive tract. However, the gut of flatworms is highly bifurcated and convoluted,
with pockets that radiate out into the deep tissue on the sides of the animal.
This is probably adaptive in this organism because it insures that no cell is
too far from the gut (and digested nutrients). Flatworms also have ameobocytes
like sponges and cnidarians, too; these transport nutrients between the endodermal
cells lining the gut and the rest of the tissues in the animal. Digestion is
both intracellular and extracellular, as in cnidarians. The major evolutionary
innovations in this group are associated with bilaterality. The nervous system
and sensory organs (eyespots and chemoreceptive auricles) are concentrated towards
the anterior end of the animal, creating a head (of sorts). Flatworms are a
diverse group, with free-living and parasitic forms. Planaria live in marine,
freshwater, and terrestrial environments. Flukes are parasitic, like the liver
flukes and blood flukes (Schistosomes) we saw in lab. Most of these species
have at least one intermediate host and a complex life cycle. At some point
in the life cycle there is usually an asexual stage in which thousands of offspring
can be produced asexually. This is particularly important for parasites that
infect different hosts in sequence, because the probability of finding new hosts
is low. Tapeworms are another group of parastic flatworms. Over 1000 species
have been described, and every vertebrate species is infected by at least one
species of tapeworm. Humans are infected by three, with sheep, pigs, cattle,
and fish acting as intermediate hosts. Tapeworms are weird. Humans ingest living
cysts in undercooked beef, lamb, pork, and fish. The immature worm hatches from
the cysts and attached to the wall of the intestine with the hooks on the scolex
- the head segment. As the worm matures and grows, it produces body segments
called proglottids. Each proglottid contains male and female reproductive organs.
As the worm creates new proglottids behind the head, older proglottids that
are farther from the head mature. The worm can be over 12m long in humans; some
grow to over 30m in length! Each proglottid is metabolically independent, absorbing
nutrients by diffusion, directly from the hosts intestinal tract. The high SA/V
ratio of these flatworms aids in this parasitic lifestyle. Each proglottid produces
egg and sperm; sperm are received by proglottids and the development of fertilized
eggs occurs in each proglottid. Eventually, the mature proglottids are shed
by the worm, and are shed from the host in feces. The proglottids are eaten
by the secondary host (sheep, pigs, cattle, and fish). The larvae hatch, bore
through the intestinal wall, and migrate to muscle tissue where they encyst.
Consumption of undercooked meat by another person completes the life cycle.
d. Phylum Annelida: Segmented worms
 Annelids
include three major groups - the polychaete worms (like we used in lab as the
outgroup for assessing shared derived traits in arthropods), earthworms (like
we dissected in lab), and leeches. Annelids inhabit marine, freshwater, and
terrestrial environments. The annelids are an ancient group, but their soft
bodies do not fossilize well. However, there are unambiguous annelids that date
to the Cambrian, and the putative polychaete Dicksonia dates from the
Vendian Period 600 mya. With the Annelida we see a fundamental change in body
plan - segmentation. Like gene duplication, the replication of body units into
separate segments easily allows for body region to specialize, or to evolve
a new function in one area without compromising the ability to perform original
functions in other segments. This redundancy is easily seen in the anterior
region of earthworms, where each segment contains a heart. Likewise, each segment
throughout the animal contains its own excretory units - nephridia. Each segment
of polychaetes has parapodia - 'side-feet' used for locomotion and respiration
(increasing the surface area for gas exchange). Annelids have a true body cavity,
they are not solid cells from the gut to the outer wall. This cavity provides
room for outpocketings of the digestive tract and other organs. In addition,
this fluid-
Annelids
include three major groups - the polychaete worms (like we used in lab as the
outgroup for assessing shared derived traits in arthropods), earthworms (like
we dissected in lab), and leeches. Annelids inhabit marine, freshwater, and
terrestrial environments. The annelids are an ancient group, but their soft
bodies do not fossilize well. However, there are unambiguous annelids that date
to the Cambrian, and the putative polychaete Dicksonia dates from the
Vendian Period 600 mya. With the Annelida we see a fundamental change in body
plan - segmentation. Like gene duplication, the replication of body units into
separate segments easily allows for body region to specialize, or to evolve
a new function in one area without compromising the ability to perform original
functions in other segments. This redundancy is easily seen in the anterior
region of earthworms, where each segment contains a heart. Likewise, each segment
throughout the animal contains its own excretory units - nephridia. Each segment
of polychaetes has parapodia - 'side-feet' used for locomotion and respiration
(increasing the surface area for gas exchange). Annelids have a true body cavity,
they are not solid cells from the gut to the outer wall. This cavity provides
room for outpocketings of the digestive tract and other organs. In addition,
this fluid- filled
space can act as a hydrostatic skeleton that allows for refined, peristaltic
locomotion. Water is incompressible, so muscles contracting in one part of the
body push the water in the coelom somewhere else, like towards the front of
the animal which extends the body forward. Annelids also have another important
difference with the other phyla we have considered - they have a digestive tract
rather than a gut cavity. In a gut cavity, new food is added and is mixed with
digesting food; there is no separation and so there can be no specialization
of digestive function. In a tract, where food enters at one end of the animal
(mouth) and passes in one direction towards the exit (anus), there can be specialization
of function along the route - creating a "dis-assembly line" that
breaks food down and harvests energy more efficiently. We see this specialization
in earthworms, where the mucular pharynx contracts to extend the mouth for ingestion,
the crop that stores food and delivers it evenly to the gizzard, which grinds
the food into smaller particles (increasing its surface area for the action
of enzymes), and the intestine where digestion and the absorption of nutrients
occurs. Earthworms eat soil, digesting the fungi, bacteria, organic molecules
and protists. They pass fecal pellets that contain the mineral components and
unabsorbed organics. By creating these pellets, they improve the ability of
water and air to travel through the soil, increasing the uptake of water and
soluble nutrients, and increasing gas exchange by plant roots. This increases
the growth rate of plants and increases the productivity of the community.
filled
space can act as a hydrostatic skeleton that allows for refined, peristaltic
locomotion. Water is incompressible, so muscles contracting in one part of the
body push the water in the coelom somewhere else, like towards the front of
the animal which extends the body forward. Annelids also have another important
difference with the other phyla we have considered - they have a digestive tract
rather than a gut cavity. In a gut cavity, new food is added and is mixed with
digesting food; there is no separation and so there can be no specialization
of digestive function. In a tract, where food enters at one end of the animal
(mouth) and passes in one direction towards the exit (anus), there can be specialization
of function along the route - creating a "dis-assembly line" that
breaks food down and harvests energy more efficiently. We see this specialization
in earthworms, where the mucular pharynx contracts to extend the mouth for ingestion,
the crop that stores food and delivers it evenly to the gizzard, which grinds
the food into smaller particles (increasing its surface area for the action
of enzymes), and the intestine where digestion and the absorption of nutrients
occurs. Earthworms eat soil, digesting the fungi, bacteria, organic molecules
and protists. They pass fecal pellets that contain the mineral components and
unabsorbed organics. By creating these pellets, they improve the ability of
water and air to travel through the soil, increasing the uptake of water and
soluble nutrients, and increasing gas exchange by plant roots. This increases
the growth rate of plants and increases the productivity of the community.
e. Phylum Mollusca: Chitons, Bivalves, Snails, and Cephalopods
 The
molluscs are also an ancient group, dating to the late pre-Cambrian Period (about
560 mya). Shelled molluscs, protected from cnidarian predators, radiate in the
Cambrian. The molluscan body plan is fairly simple, best seen in the most primitive
extant lineage, the Polyplacophorans (Chitons). They are bilaterally symmetrical
with a complete digestive tract. Their body is covered by a hood-like structure
called the mantle. This mantle creates a pocket between the body of the animal
and the hood. Gills are present in this mantle cavity. The bottom of the animal
is a muscular foot. In many molluscs, the mantle secretes a shell. In the polyplacophorans,
the shell is composed of 8 units or plates, suggesting the segmented ancestry
of the group. However, more derived molluscs have lost most indications of segmentation.
Although molluscs take a wide variety of shapes, from snails to clams to squid,
all of these shapes are just variations on this basic theme.
The
molluscs are also an ancient group, dating to the late pre-Cambrian Period (about
560 mya). Shelled molluscs, protected from cnidarian predators, radiate in the
Cambrian. The molluscan body plan is fairly simple, best seen in the most primitive
extant lineage, the Polyplacophorans (Chitons). They are bilaterally symmetrical
with a complete digestive tract. Their body is covered by a hood-like structure
called the mantle. This mantle creates a pocket between the body of the animal
and the hood. Gills are present in this mantle cavity. The bottom of the animal
is a muscular foot. In many molluscs, the mantle secretes a shell. In the polyplacophorans,
the shell is composed of 8 units or plates, suggesting the segmented ancestry
of the group. However, more derived molluscs have lost most indications of segmentation.
Although molluscs take a wide variety of shapes, from snails to clams to squid,
all of these shapes are just variations on this basic theme.
 Snails
have put a 'twist' on molluscan evolution. The shell and mantle rotate during
development, placing the mantle cavity at the back of the animal. Many snails
are grazers; they scrape algae off rocks with a file-like tongue called a radula.There
are many predatory snails, however. Oyster drills have a tough radula, and they
scrape a hole through oyster shells. They then extend their proboscis through
the hole and consume the oyster. Coneshells, in the genus Conus, have
a modified radula that takes the shape and function of a harpoon. They stab
fish and inject a potent toxin; some Conus snail toxins are also lethal
to humans. Almost all molluscs are marine or aquatic; only some snails have
colonized land.The most coloful molluscs are nudibranchs (sea slugs); they prey
on hydroids and coral polyps, and some even prey on venomous jellyfish. Some
species are able to digest the cnidarians without disturbing their cnidocytes,
and these cells are moved to the back of the nudibranch and protect it against
the nudibranch's own predators. The bright colors are though to be warning coloration.
The protuberances on the back are the "branch's" - the external gill-like
respiratory organs.
Snails
have put a 'twist' on molluscan evolution. The shell and mantle rotate during
development, placing the mantle cavity at the back of the animal. Many snails
are grazers; they scrape algae off rocks with a file-like tongue called a radula.There
are many predatory snails, however. Oyster drills have a tough radula, and they
scrape a hole through oyster shells. They then extend their proboscis through
the hole and consume the oyster. Coneshells, in the genus Conus, have
a modified radula that takes the shape and function of a harpoon. They stab
fish and inject a potent toxin; some Conus snail toxins are also lethal
to humans. Almost all molluscs are marine or aquatic; only some snails have
colonized land.The most coloful molluscs are nudibranchs (sea slugs); they prey
on hydroids and coral polyps, and some even prey on venomous jellyfish. Some
species are able to digest the cnidarians without disturbing their cnidocytes,
and these cells are moved to the back of the nudibranch and protect it against
the nudibranch's own predators. The bright colors are though to be warning coloration.
The protuberances on the back are the "branch's" - the external gill-like
respiratory organs.
 Bivalves
are an unusual group of molluscs. Bivalves are aquatic/marine filter feeders;
they lay on the sediment and filter water between the 'valves' of their shell.
They are bilaterally symmetrical; the midline of their back runs along the hinge
of their shell - so their shell comes down on both sides of the animal, covering
the animal and the foot. However, since the animals are sessile filter feeders,
their nervous system has evolved to be decephalized (decentralized). They don't
have a head - rather, because they do not move, their sensory systems have been
redistributed around the organism so they can perceive their environment from
all directions. Scallops, for instance, have eyes all around the margin of their
mantle and shell. So again, here is a sort of exception that proves the rule:
bilaterality directs a pattern of motion, which selects for cephalization at
one end. Some sessile organisms that don't move through the environment are
decephalized.
Bivalves
are an unusual group of molluscs. Bivalves are aquatic/marine filter feeders;
they lay on the sediment and filter water between the 'valves' of their shell.
They are bilaterally symmetrical; the midline of their back runs along the hinge
of their shell - so their shell comes down on both sides of the animal, covering
the animal and the foot. However, since the animals are sessile filter feeders,
their nervous system has evolved to be decephalized (decentralized). They don't
have a head - rather, because they do not move, their sensory systems have been
redistributed around the organism so they can perceive their environment from
all directions. Scallops, for instance, have eyes all around the margin of their
mantle and shell. So again, here is a sort of exception that proves the rule:
bilaterality directs a pattern of motion, which selects for cephalization at
one end. Some sessile organisms that don't move through the environment are
decephalized.
 The
last group of molluscs are the other-wordly cephalopods. Although they look
very different from a chiton, the evolution of their body plan entailed a rather
simple modification - the folding of the animal head to tail, and the elongation
of the back. The folding of the animal created an anterior half to the foot,
which evolved into tentacles, and a posterior half to the foot, which evolved
into the siphon. The mantle forms a muscular cowl over the organism, and water
can be sucked into the mantle cavity through the siphon, aerating the gills,
and then expelled by the muscular contraction of the mantle - propelling the
animal in a direction determined by the aiming of the siphon. As their name
implies, the cephalopods have a big head - and it is a brainy, smart head, too.
This correlates with their predatory behavior. As a general rule, predators
are pretty clever animals - at least more clever than their prey. This is especially
true for highly active and mobile predators like cephalopods, cats, and dogs.
Cephalopods are the largest living invertebrates, and they are also generally
regarded as the smartest. They can learn by mimicry. In a famous study, two
octopuses were placed in neighboring glass aquaria that had red and blue balls
at one end. One octopus had been trained to travel to the other end of the aquarium
and pick up a red ball to receive food. After observing this behavior, the second
untrained octopus would travel to the end of its aquarium and pick up a red
ball (not a blue one). That is alot of higher processing occuring. Within the
cephalopods, the evolutionary trend has been the reduction, internalization,
and loss of the shell. A shell is very heavy; reducing its size increases the
mobility of the predator. Internalizing the shell, as a rigid strut ("pen")
in squid allows it to help the hydrodynamics of the organism. For octopus,which
are sit and wait predators, the loss of the shell was adaptive because it meant
that octopus could squeeze into very narrow cavities to escape their predators
and to surprize their prey.
The
last group of molluscs are the other-wordly cephalopods. Although they look
very different from a chiton, the evolution of their body plan entailed a rather
simple modification - the folding of the animal head to tail, and the elongation
of the back. The folding of the animal created an anterior half to the foot,
which evolved into tentacles, and a posterior half to the foot, which evolved
into the siphon. The mantle forms a muscular cowl over the organism, and water
can be sucked into the mantle cavity through the siphon, aerating the gills,
and then expelled by the muscular contraction of the mantle - propelling the
animal in a direction determined by the aiming of the siphon. As their name
implies, the cephalopods have a big head - and it is a brainy, smart head, too.
This correlates with their predatory behavior. As a general rule, predators
are pretty clever animals - at least more clever than their prey. This is especially
true for highly active and mobile predators like cephalopods, cats, and dogs.
Cephalopods are the largest living invertebrates, and they are also generally
regarded as the smartest. They can learn by mimicry. In a famous study, two
octopuses were placed in neighboring glass aquaria that had red and blue balls
at one end. One octopus had been trained to travel to the other end of the aquarium
and pick up a red ball to receive food. After observing this behavior, the second
untrained octopus would travel to the end of its aquarium and pick up a red
ball (not a blue one). That is alot of higher processing occuring. Within the
cephalopods, the evolutionary trend has been the reduction, internalization,
and loss of the shell. A shell is very heavy; reducing its size increases the
mobility of the predator. Internalizing the shell, as a rigid strut ("pen")
in squid allows it to help the hydrodynamics of the organism. For octopus,which
are sit and wait predators, the loss of the shell was adaptive because it meant
that octopus could squeeze into very narrow cavities to escape their predators
and to surprize their prey.
Protostomes: Ecdysozoans
 f.
Phylum Nematoda: Roundworms
f.
Phylum Nematoda: Roundworms
Like all Ecdysozoans, nematodes have a cuticle that must be shed in order for the animal to grow. In most nematodes, the cuticle is shed four times during the growth phase of the organism. Nematodes have a complete digestive tract and some cephalization, but they generally have a somewhat simplied and reduced body plan. Many are free-living in soil and aquatic environments, but many are parasitic, infecting all other forms of multicellular life. Nematode infections in humans cause trichinosis, filariasis, elephantiasis, and ascariasis.
g. Phylum Tardigrada: Waterbears
This is a curious phylum of the toughest animals on earth. They can tolerate temperatures above 150 C for short periods, and can be dehydrated to 1% of their normal water content for 10 years and be rehydrated. They can also tolerate the vaccuum of space and 6000 atmospheres of pressure! They are found in terrestrial, marine, and aquatic habitats from the poles to the tropics. Most eat plants, algae, or bacteria. With the onychophorans, they are the sister group to the arthropods.
h. Phylum Onychophora: Velvetworms
These animals have segmented bodies but lobe-like legs. They are thought to be a sister group to the arthropods, perhaps representing the living discendants of the link between the annelids and arthropods. They are tropical, and they capture prey by squirting them with a glue to subdue them.
i. Phylum Arthropoda
 The
arthropods are the most diverse animal group on earth, accounting for 85% of
all animal species. The are an ancient group that came to dominate during the
Cambrian period. The earliest groups were the Chelicerates and Trilobites. The
chelicerates are identified by having only two appendages forward of the mouth
- these are the small, typically pincer-like chelicerae. In spiders, the chelicerae
have evolved into fangs that deliver poison to subdue prey. The other group
of chelicerates are the Horseshoe crabs - an ancient relictual taxon. The other
major clade of arthropods are the mandibulata, which divide into the myriapods
(millipedes and centipedes) and the crustacea. Myriapods are segmented, without
much specialization; each body segment has similar paired appendages used for
walking Centipedes have one pair of legs per segment, and they are predatory;
they have poison glands like spiders and inject their prey to immobilize them.
Millipedes have two apirs of legs per segment. they are slower than centipedes,
and they feed on detritus. In the Eucrustacea, the segments have paired appendages
but they differ in length and function. In addition, they are biramous - having
two roots. One root is a leg, and the other is a gill. Curiously, the insects
are a group derived from aquatic crustaceans. They have flourished and radiated
in terrestrial environments like no other group in the history of our planet.
They have reduced, fused body parts, with appendages limited to head and thoracic
segments. Insect diversity is staggering, and this diversity has interested
ecologist for along time. Why are there so many species of insects? Well, their
diversity is testimony to their adaptability - their exoskeleton means that
they can tolerate dessication. Their small size means that they don't need many
resources to survive. Their power of flight means that they can disperse easily.
This means that they can find resources, but it also means that they are prone
to geographic isolation (and speciation). Finally, they are fecund - most insects
can lay many eggs and thus establish large populations quickly. For all these
reasons and many more, insects are prone to succeed and speciate.
The
arthropods are the most diverse animal group on earth, accounting for 85% of
all animal species. The are an ancient group that came to dominate during the
Cambrian period. The earliest groups were the Chelicerates and Trilobites. The
chelicerates are identified by having only two appendages forward of the mouth
- these are the small, typically pincer-like chelicerae. In spiders, the chelicerae
have evolved into fangs that deliver poison to subdue prey. The other group
of chelicerates are the Horseshoe crabs - an ancient relictual taxon. The other
major clade of arthropods are the mandibulata, which divide into the myriapods
(millipedes and centipedes) and the crustacea. Myriapods are segmented, without
much specialization; each body segment has similar paired appendages used for
walking Centipedes have one pair of legs per segment, and they are predatory;
they have poison glands like spiders and inject their prey to immobilize them.
Millipedes have two apirs of legs per segment. they are slower than centipedes,
and they feed on detritus. In the Eucrustacea, the segments have paired appendages
but they differ in length and function. In addition, they are biramous - having
two roots. One root is a leg, and the other is a gill. Curiously, the insects
are a group derived from aquatic crustaceans. They have flourished and radiated
in terrestrial environments like no other group in the history of our planet.
They have reduced, fused body parts, with appendages limited to head and thoracic
segments. Insect diversity is staggering, and this diversity has interested
ecologist for along time. Why are there so many species of insects? Well, their
diversity is testimony to their adaptability - their exoskeleton means that
they can tolerate dessication. Their small size means that they don't need many
resources to survive. Their power of flight means that they can disperse easily.
This means that they can find resources, but it also means that they are prone
to geographic isolation (and speciation). Finally, they are fecund - most insects
can lay many eggs and thus establish large populations quickly. For all these
reasons and many more, insects are prone to succeed and speciate.
In looking at the mandibulate groups, we see a nice example of an important evolutionary pattern: replication, specialization, and reduction. The myriapods have probably preserved the ancestral body plan within the Arthropoda. Like their probable annelid ancestors, the myriapods are composed of many similar segments. The Eucrustacea represent the next step in the evolution of many body plans - specialization of different body segments for particular functions. So, although every segment of the body of a eucrustacean has paired appendages (like myriapods), those appendages are different lengths and sizes and perform different functions. Appendages in the head are antennae and tiny feeding pincers. Appendages in the thorax are legs and claws (like in lobsters and crabs). Appendages in the abdomen are involved in reproduction (holding eggs, for instance). Next time you have lobster, crab, or shrimp, take a closer look at your meal and notice the paired appendages on each body sement that have become specialized for different functions. In the most derived group of crustaceans - the insects - there is fusion of segments in a given body region and the elimination of redundancy. The abdominal segments have fused into one body region, and the appendages have been lost. The thoracic segments have fused into one body region, and most segments have lost their legs - reducing the redundancy and getting by with 6. Likewise, head segments have fused, and insects have only one pair of antennae (unlike the 2 pairs of eucrustaceans). So, in this one clade, we see the important pattern of segmentation, specialization, and fusion / reduction as a mechanism to produce evolutionary novelty.
Deuterostomes:
j. Phylum Echinodermata: Sea stars, sea urchins, sea cucumbers, sand dollars
 Like
all deuterstomes, the blastopore forms the anus in this phylum. The echinoderms
have an internal skeleton consisting of separate plates that interlock. This
allows them flexibility, but also remarkable rigidity. The body axis of a starfish
is from the bottom, called the oral surface, to the top, called the aboral surface.
So, the legs radiate from the 'waist' of the animal. This symmetry is called
'pentaramous' symmetry. The other interesting adaptiation of echinoderms are
the internal canal system. They have tubes that fill tube feet, causing them
to extend and retract as they fill and deflate. This internal system of canals
is continuous with the environment, with a valve called the madreporite that
regulates water flow from the environment to the canals. The digestive tract
of a starfish is short, from the oral to aboral surface. But the efficiency
is increased by having digestive glands radiate out into the arms or the starfish.
Gonads are also in the limbs.
Like
all deuterstomes, the blastopore forms the anus in this phylum. The echinoderms
have an internal skeleton consisting of separate plates that interlock. This
allows them flexibility, but also remarkable rigidity. The body axis of a starfish
is from the bottom, called the oral surface, to the top, called the aboral surface.
So, the legs radiate from the 'waist' of the animal. This symmetry is called
'pentaramous' symmetry. The other interesting adaptiation of echinoderms are
the internal canal system. They have tubes that fill tube feet, causing them
to extend and retract as they fill and deflate. This internal system of canals
is continuous with the environment, with a valve called the madreporite that
regulates water flow from the environment to the canals. The digestive tract
of a starfish is short, from the oral to aboral surface. But the efficiency
is increased by having digestive glands radiate out into the arms or the starfish.
Gonads are also in the limbs.
k. Phylum Hemichordata: Acorn Worms
The hemichordates are an exclusively marine group. There are interesting because they have two of the four characteristics seen in the chordate: they have pharygeal gills slits and they have a hollow dorsal nerve chord. If you remember from the earthworm dissection, other phyla have ventral nerve chords.There are two major groups of hemichordates, and one group forms protective tubes out of collagen they secrete, and they filter feed with echinoderm-like arms.
l. Phylum Chordata: Tunicates, Cephalochordates, and Vertebrates
All chordates have four characteristics at some point in their development: 1) pharyngeal folds (or gill slits), 2) hollow dorsal nerve tube, 3) notochord, and 4) a post-anal tail. As you might suspect, the similarities among organisms in this phylum are most evident in the larval stages. Cephalochordates like Amphioxus probably maintain the ancestral condition within the phylum. They have swimming larvae, but adults burrow and either feed on detritus or filter feed using their gills as a sieve. The oldest chordate fossils - Pikaia from the Cambrian Period -have a morphology very similar to modern cephalochordates. The Urochordata - the tunicates - have adapted to a more specialized filter-feeding lifestyle. Although their larvae are mobile and similar to Amphioxus, they settle out on a substrate, on their backs, and fold head-to-tail. The pharynx becomes a filtering 'basket'.
 |
 |
 |
Urochordata: Tunicates |
Cephalochordata: Amphioxus |
Vertebrata |
Vertebrates represent the evolutionary lineage that went in the other direction: not towards a sedentary lifestyle but towards an active lifestyle. For the remainder of this unit, we will look at the evolutionary radiations within the vertebrates. We will see a typical pattern throughout this unit. A new group will evolve a novel trait or way of life. This will allow it to use a new environment, or use the environment in a new way. These lifestyles are called "adaptive zones", and they relate to ecological niches. Creation or entry into a new niche or zone means that there is no competition for resources. Under these conditions, almost any strategy using this new novel innovation will work - typically we see a radiation of new species exploiting this new way of life. As this adaptive zone or niche fills with species, they begin to compete. This leads to 'competitive contraction' of diversity, with only a few winners. The same thing happens in any situation where there are potentially limted resources. A new innovation, like the internet, provides opportunities to new businesses. Many flourish initially - like the many browser providers that were present in the '90's. Anyone remember when Netscape was the primary internet provider? Economists call that period the "dot-com boom", when internet companies could get a start just by colonizing this new open marketplace. As companies grew, they began to compete; and there were winners that maintained their position in the niche, and losers that went extinct. The same thing can happen in ecological systems. Competitive contraction can occur within a clade, or as a consequence of a more efficient, new evolutionary novelty arising. Henry Ford's Model T not only put many other car makers out of business, it also puts LOTS of horse-driven carriage companies out of business! So, as a more advanced, efficient group evolves, we sometimes see contraction in the abundance of ancestral forms... just as Darwin predicted.
 The
jawless fishes:
The vertebrates are distinguished by having an internal supportive spine that
is subdivided into units called vertebrae. This segmentation of the vertebral
column allows the animal to bend and move with more precision; with some parts
of the body moving while other parts remain stationary. The most primitive members
of this clade are the jawless fishes. The modern representatives are the hagfish
and the lampreys. The hagfish actually lack a subdivided vertebral column, but
they do have a skull surrounding their brain. As such, sometimes the "vertebrata"
are more specifically described as having a skull (to include the hagfish).
Both hagfish and lampreys have immature stages that look almost identical to
cephalochordates, also suggesting their primitive position within this clade.
So, the jawless fish evolve in the Cambrian and radiate in the Ordovician and
Silurian Periods, with many large, detritivorous and filter feeding species
scouring the benthos for food. In more advanced forms, the mouth cavity and
pharynx evolved for feeding, relieving the gills of this filtering function.
These jawless fish probably sucked in small invertebrates by rapidly opening
their mouth.
The
jawless fishes:
The vertebrates are distinguished by having an internal supportive spine that
is subdivided into units called vertebrae. This segmentation of the vertebral
column allows the animal to bend and move with more precision; with some parts
of the body moving while other parts remain stationary. The most primitive members
of this clade are the jawless fishes. The modern representatives are the hagfish
and the lampreys. The hagfish actually lack a subdivided vertebral column, but
they do have a skull surrounding their brain. As such, sometimes the "vertebrata"
are more specifically described as having a skull (to include the hagfish).
Both hagfish and lampreys have immature stages that look almost identical to
cephalochordates, also suggesting their primitive position within this clade.
So, the jawless fish evolve in the Cambrian and radiate in the Ordovician and
Silurian Periods, with many large, detritivorous and filter feeding species
scouring the benthos for food. In more advanced forms, the mouth cavity and
pharynx evolved for feeding, relieving the gills of this filtering function.
These jawless fish probably sucked in small invertebrates by rapidly opening
their mouth.
 The
Gnathostomes: The Jawed Vertebrates: Jawed fishes evolved
in the Silurian and came to dominate and radiate in the Devonian - the "Age
of Fishes". Jaws evolved from the anterior gill arches, which flex and
became associated withe the mouth. Jaws allowed fish to become more efficient
predators, killing things bigger than could fit in their mouth through suction,
or strong enough to escape the suctioning force. In short, jaws were adaptive
because fish now became mobile, active predators - entering a niche previously
only occupied by large invertebrate predators. The first jawed fish radiation
were the placoderms, symbolized by the frightening Arthrodire predators like
Dunkleosteus, with its huge slicing jaw plates. The placoderms were
very heavy, however, and were displaced by more efficient swimmers - the cartilaginous
jawed fishes (Class Chondrichthyes). Modern representatives of this group are
sharks and rays.
The
Gnathostomes: The Jawed Vertebrates: Jawed fishes evolved
in the Silurian and came to dominate and radiate in the Devonian - the "Age
of Fishes". Jaws evolved from the anterior gill arches, which flex and
became associated withe the mouth. Jaws allowed fish to become more efficient
predators, killing things bigger than could fit in their mouth through suction,
or strong enough to escape the suctioning force. In short, jaws were adaptive
because fish now became mobile, active predators - entering a niche previously
only occupied by large invertebrate predators. The first jawed fish radiation
were the placoderms, symbolized by the frightening Arthrodire predators like
Dunkleosteus, with its huge slicing jaw plates. The placoderms were
very heavy, however, and were displaced by more efficient swimmers - the cartilaginous
jawed fishes (Class Chondrichthyes). Modern representatives of this group are
sharks and rays.
 The
Devonian also saw the radiation of the most successful group of fish on earth,
the bony fish (Class Osteichthyes). Unlike cartilage, bone is hollow and light;
it is also more rigid than cartilage, providing more resistance and efficiency
to muscles. The bony fish also had a swim bladder, whcih they used to maintain
neutral bouyancy in the water column. This means that they can maintain position
without swimming; Condrichthyes must swim to maintain vertical position in the
water column - otherwise they sink. These two adaptations of the bony fishes
made them more energetically efficient and faster than the cartilaginous fish,
and the bony fishes have radiated to the point where they represent 40% of all
vertebrate species today (not just fish - vertebrates..!!). Two important clades
evolved within the bony fishes: 1) the ray-finned fishes that came to dominate
the aquatic environments of the planet; and 2) the lob-finned fishes that radiated
into shallow-water environments. The lobe-finned fishes, swimming in shallow,
oxygen-poor waters of the Devonian, gulping air at the surface and pulling themselves
along on their forefins, evolved into the terrestrial tetrapods. Arthropods
had already colonized land, so there was ample food and oxygen for a large terrestrial
predator. As you have seen, the fossil record nicely documents the transition
from fish to these primitive amphibians.
The
Devonian also saw the radiation of the most successful group of fish on earth,
the bony fish (Class Osteichthyes). Unlike cartilage, bone is hollow and light;
it is also more rigid than cartilage, providing more resistance and efficiency
to muscles. The bony fish also had a swim bladder, whcih they used to maintain
neutral bouyancy in the water column. This means that they can maintain position
without swimming; Condrichthyes must swim to maintain vertical position in the
water column - otherwise they sink. These two adaptations of the bony fishes
made them more energetically efficient and faster than the cartilaginous fish,
and the bony fishes have radiated to the point where they represent 40% of all
vertebrate species today (not just fish - vertebrates..!!). Two important clades
evolved within the bony fishes: 1) the ray-finned fishes that came to dominate
the aquatic environments of the planet; and 2) the lob-finned fishes that radiated
into shallow-water environments. The lobe-finned fishes, swimming in shallow,
oxygen-poor waters of the Devonian, gulping air at the surface and pulling themselves
along on their forefins, evolved into the terrestrial tetrapods. Arthropods
had already colonized land, so there was ample food and oxygen for a large terrestrial
predator. As you have seen, the fossil record nicely documents the transition
from fish to these primitive amphibians.
 The
Tetrapods: The
tetrapod clade includes all vertebrates descended from the first animals to
walk on land. The radiation of early tetrapods is very impressive; the adaptive
zone of the Carboniferous swamps was very large and well-suited to amphibian
animals reliant on water to lay their eggs. In addition, the huge amount of
biological productivity (lots of photosynthesis and less decompostion, remember?)
led to large populations of herbivorous insects that were eay prey for large
land vertebrates. The Carboniferous could well be called the "Age of Amphibians".
Many groups of early 'amphibians' radiate during this period, collectively called
the 'stem tetrapods'. However, the dry periods of the Permian and Mesozoic were
not optimal for these animals limited to areas near water. Today, three groups
of amphibians remain. The Caecilians are unusual, rare, legless amphibians.
Salamanders are the second group, and the giant Japanese salamander is the largest
living amphibian - reaching six feet in length! The most diverse group is the
Anura - mening "tailless" - the frogs and toads. Most amphibians have
gills or lungs at some point in their life cycle, but many also (or exclusively)
respire through their skin. For this to happen, the gases must diffuse into
a thin film of water, and then move across cell membranes by diffusion. So,
most amphibians need to keep their skin moist to respire; and it also means
that their skin, as an absorptive surface, is also prone to absorb toxins in
the environment. For these reasons, amphibians are particularly sensitive to
pollutants and environmental contaminants. Over the last 50 years, there has
been a dramatic decline in amphibian populations worldwide, probably due to
exposure to human-produced pollution, climate change, and chytrid fungal infections.
It is tough to say which of these variables is 'most' responsible for the decline.
In all likelihood, each factor increases sensitivity to the other two. In any
case, these three factors are having a decimating effect on amphibian populations.
The
Tetrapods: The
tetrapod clade includes all vertebrates descended from the first animals to
walk on land. The radiation of early tetrapods is very impressive; the adaptive
zone of the Carboniferous swamps was very large and well-suited to amphibian
animals reliant on water to lay their eggs. In addition, the huge amount of
biological productivity (lots of photosynthesis and less decompostion, remember?)
led to large populations of herbivorous insects that were eay prey for large
land vertebrates. The Carboniferous could well be called the "Age of Amphibians".
Many groups of early 'amphibians' radiate during this period, collectively called
the 'stem tetrapods'. However, the dry periods of the Permian and Mesozoic were
not optimal for these animals limited to areas near water. Today, three groups
of amphibians remain. The Caecilians are unusual, rare, legless amphibians.
Salamanders are the second group, and the giant Japanese salamander is the largest
living amphibian - reaching six feet in length! The most diverse group is the
Anura - mening "tailless" - the frogs and toads. Most amphibians have
gills or lungs at some point in their life cycle, but many also (or exclusively)
respire through their skin. For this to happen, the gases must diffuse into
a thin film of water, and then move across cell membranes by diffusion. So,
most amphibians need to keep their skin moist to respire; and it also means
that their skin, as an absorptive surface, is also prone to absorb toxins in
the environment. For these reasons, amphibians are particularly sensitive to
pollutants and environmental contaminants. Over the last 50 years, there has
been a dramatic decline in amphibian populations worldwide, probably due to
exposure to human-produced pollution, climate change, and chytrid fungal infections.
It is tough to say which of these variables is 'most' responsible for the decline.
In all likelihood, each factor increases sensitivity to the other two. In any
case, these three factors are having a decimating effect on amphibian populations.
The Amniota: Within the tetrapods, a new type of animal evolved in the Carboniferous; an animal that laid an egg surrounded by a series of membranes and a shell. These layers served to protect the developing embryo and yolk sac from dessication, allowing these species to colonize land further from water and exploit food resources and habitats that amphibians could not reach. This clade of animals is the amniotes, and their evolutionary innovation is the amniotic egg. The primitive condition within this group probably produced a leathery covering, much like turtles. A more rigid, desiccation resistant egg like birds have is probably a derived trait, as is the retention of the egg and live bearing of young, as seen in some snakes, and most mammals. The amniotes radiated during the Permian Period, when the drying of the unified land mass of Pangaea gave them a decided advantage over the ancestral, amphibious tetrapods. Three groups evolved at this time, distinguished by their skull morphology.
 The
Synapsids and the Mammals: The first group to dominate
were the synapsids. They dominated during the Permian, and were represented
by the great "sail-finned" Pelycosaurs and the Therapsid lineages
(the Gorgonopsians, the Dicynodonts, and the Cynodonts). The synapsid lineage
that is alive today are the Mammals. The transition from ancestral to derive
synapsids is well preserved in the fossil record, documenting the evolution
of the inner ear from the ancestral jaw bones, and the evolution of complex
and specialized dentition. The first true mammals are the Morganucodonts that
evolved in the Jurassic Period of the Mesozoic, about 200 mya. Ancestral synapsids
were large carnivores that dominated the landscape; a niche that would be filled
by the diapsid dinosaurs during the Mesozoic. The mammals were small, noctural
insectivores and scavengers. They radiated into these niches in the Jurassic,
perhaps because their endothermy allowed them to use the cool night more effectively
and under cover of darkness from larger predators. The surviving lineage of
this radiation are the three species of monotremes alive today: the platypus
and two species of echidna. These mammals lay eggs. Their offspring hatch, and
then nuzzle the belly of their mothers. Their mothers have sweat glands that
have evolved to produce a nutrient rich milk - they are mammary glands. However,
even the modern representatives lack nipples on these glands - so they "sweat"
milk onto their belly and the the hatchlings lick it up. Placental and marsupial
mammals evolved later, in the Cretaceous Period. Marsupials do not lay eggs;
they give live birth to a very small, embryonic newborn. The newborn climbs
from the genital opening to the pouch - a flap of skin covering the nipples
of the mammary glands. The embryo attaches to a nipple and feeds nearly continuously,
completing development in the pouch. This allows the mother to carry the embryo
with her for a longer period; there are no eggs that must be left periodically
for the mother to feed. The last group of mammals, the placentals, also retain
the embryo and give live birth. However, they retain the embryo much longer,
often until the offspring is capable of independent activity. The efficient
feeding of the embryo is facilitated by the placenta - which allows nutrients
in the bloodstream of the mother to pass directly into the bloodstream of the
offspring. This is far more efficient, energetically, than the mother turning
the nutrients into milk, that must be consumed and digested by the offspring.
So, placental mammals can develop more rapidly than marsupials. Some placentals
produce offspring that can walk immediately; these are precocial young. Other
species, like cats and humans, produce offspring that still need considerable
parental care. These offspring are called altricial.
The
Synapsids and the Mammals: The first group to dominate
were the synapsids. They dominated during the Permian, and were represented
by the great "sail-finned" Pelycosaurs and the Therapsid lineages
(the Gorgonopsians, the Dicynodonts, and the Cynodonts). The synapsid lineage
that is alive today are the Mammals. The transition from ancestral to derive
synapsids is well preserved in the fossil record, documenting the evolution
of the inner ear from the ancestral jaw bones, and the evolution of complex
and specialized dentition. The first true mammals are the Morganucodonts that
evolved in the Jurassic Period of the Mesozoic, about 200 mya. Ancestral synapsids
were large carnivores that dominated the landscape; a niche that would be filled
by the diapsid dinosaurs during the Mesozoic. The mammals were small, noctural
insectivores and scavengers. They radiated into these niches in the Jurassic,
perhaps because their endothermy allowed them to use the cool night more effectively
and under cover of darkness from larger predators. The surviving lineage of
this radiation are the three species of monotremes alive today: the platypus
and two species of echidna. These mammals lay eggs. Their offspring hatch, and
then nuzzle the belly of their mothers. Their mothers have sweat glands that
have evolved to produce a nutrient rich milk - they are mammary glands. However,
even the modern representatives lack nipples on these glands - so they "sweat"
milk onto their belly and the the hatchlings lick it up. Placental and marsupial
mammals evolved later, in the Cretaceous Period. Marsupials do not lay eggs;
they give live birth to a very small, embryonic newborn. The newborn climbs
from the genital opening to the pouch - a flap of skin covering the nipples
of the mammary glands. The embryo attaches to a nipple and feeds nearly continuously,
completing development in the pouch. This allows the mother to carry the embryo
with her for a longer period; there are no eggs that must be left periodically
for the mother to feed. The last group of mammals, the placentals, also retain
the embryo and give live birth. However, they retain the embryo much longer,
often until the offspring is capable of independent activity. The efficient
feeding of the embryo is facilitated by the placenta - which allows nutrients
in the bloodstream of the mother to pass directly into the bloodstream of the
offspring. This is far more efficient, energetically, than the mother turning
the nutrients into milk, that must be consumed and digested by the offspring.
So, placental mammals can develop more rapidly than marsupials. Some placentals
produce offspring that can walk immediately; these are precocial young. Other
species, like cats and humans, produce offspring that still need considerable
parental care. These offspring are called altricial.
The Diapsid Reptiles: The other major group of amniotes are the diapsids. They radiated into two major groups, the lepidosaurs that walked with their legs splayed out and the archosaurs that walked with their legs underneath.
 The
archosaurs would dominate during the Mesozoic Era, with the great
radations of the Crocodylomorphs, Pterosaurs, and Dinosaurs. Two groups of Archosaurs
persists today - the crocodilians (caimen, gavials, alligators, and crocodiles),
and the birds. Although several groups of dinosaurs evolved feathers - probably
first for attracting mates or communicating, and then for insulation - only
in the birds did these feathers evolve for powered flight. Associated shared
derived characters are light, hollow, fused bones, the fusion of digits in the
forelimbss, a keeled sternum for anchoring large breast muscles used for flight,
and a 'wishbone' - a union of the two clavicles that acts like a spring during
the flight stroke. The loss of teeth is also a derived trait in this group,
as is the evolution of an interesting respiratory system of air sacs. Birds
have a one way lung. Air that's inhaled goes to a posterior air sac. On the
first exhalation, air is moved forward through the lung. The second inhalation
refills the posterior air sac, and the next exhalation pulls deoxygenated air
forward into the anterior air sac and out of the animal. This modification,
like a one-way digestive tract, improves the efficiency of the organ. Now, the
air in the lung is always oxygenated; it is not mixed with deoxygenated air
like in the sac-like lungs of other animals. It's not surprizing that birds
would evolve this system, as the metabolic demands of flight are much more significant
than other forms of locomotion.
The
archosaurs would dominate during the Mesozoic Era, with the great
radations of the Crocodylomorphs, Pterosaurs, and Dinosaurs. Two groups of Archosaurs
persists today - the crocodilians (caimen, gavials, alligators, and crocodiles),
and the birds. Although several groups of dinosaurs evolved feathers - probably
first for attracting mates or communicating, and then for insulation - only
in the birds did these feathers evolve for powered flight. Associated shared
derived characters are light, hollow, fused bones, the fusion of digits in the
forelimbss, a keeled sternum for anchoring large breast muscles used for flight,
and a 'wishbone' - a union of the two clavicles that acts like a spring during
the flight stroke. The loss of teeth is also a derived trait in this group,
as is the evolution of an interesting respiratory system of air sacs. Birds
have a one way lung. Air that's inhaled goes to a posterior air sac. On the
first exhalation, air is moved forward through the lung. The second inhalation
refills the posterior air sac, and the next exhalation pulls deoxygenated air
forward into the anterior air sac and out of the animal. This modification,
like a one-way digestive tract, improves the efficiency of the organ. Now, the
air in the lung is always oxygenated; it is not mixed with deoxygenated air
like in the sac-like lungs of other animals. It's not surprizing that birds
would evolve this system, as the metabolic demands of flight are much more significant
than other forms of locomotion.
The Lepidosaurs also radiated during the Mesozoic, but did not do so as dramatically as the Archosaurs. Today, the surviving representatives are the Squamata (lizards and the derived snakes) and their sister group, the Sphenodonts with two surviving relict species of Tuatara native to New Zealand. The tuatara maintains the most primitive amniote characteristics, and so is very valuable in studying the phylogeny of diapsid reptiles. The derived squamates, the modern lizards and snakes, have scales, a double hinged jaw (particularly important in snakes), and a hinge on the top of their skull.
That concludes a look at the phyla of animals. Next we will take a brief look at the evolutionary history of our own small twig on the mammalian limb.
Study Questions:
1. What is an amniotic egg and what benefit does it provide? Why did amniotes radiate in the Permian?
2. Name two diseases that nematodes cause.
3. List the characteristics of the Chordates
4. How did the jaw evolve and what benefit did it provide?
5. What is the benefit of a highly convoluted gut in flatworms?
6. What is the advantage of a digestive tract, compared to a gut sac?
7. Why was segmentation - the replication of body units - such an important evolutionary innovation? (Think of the advantages of gene duplication.)
8. Why have clams "lost their head"?
9. What are the stages of animal embryogenesis?
10. Distinguish "protostome" and "deuterostome".
11. Why are "goofy" characteristics like whether the blastopore forms the mouth or anus used to reconstruct phylogenies? Explain in terms of the three reasons why organisms might share a characteristic.
12. What are the two body types seen in Cnidarians?
13. Why is bilateral symmetry, rather than radial symmetry, associated with "cephalization" in motile organisms?
14. How do showy flowers increase the efficiency of pollen transfer?
15. What is the adaptive advantage of fleshy fruit production?
16. The two major innovations represented in the gymnosperms are seeds and pollen. Explain what each of these structures IS (in the contex of a plant life cycle), and explain how each structure allowed plants to colonize dry habitats.
17. What challenges face an aquatic algae as it tries to acquire light, gases, and nutrients?
18. To a photosynthetic organism, what are the benefits of colonizing land? What is the major "down side"?
19. Why can vascular plants be tall while non-vascular plants are short.
20. How do fungi acquire energy?
21. Fungi are primary decomposers. What other group of organisms are decomposers? What chemicals do these groups produce to kill one another (we use these chemicals to kill bacteria and fungi, too!)
22. Describe the endosymbiotic theory of organelle origin, and provide evidence for this theory.
23. What are the three domains of life?
24. Why are archaeans and bacteria so much more genetically diverse than plants, animals, and fungi?
25. All eukaryotes are aerobic. List four types of metabolism that prokaryotes have with respect to oxygen.