 I.
A Brief History of LifeI.
A Brief History of Life
I.
A Brief History of LifeI.
A Brief History of Life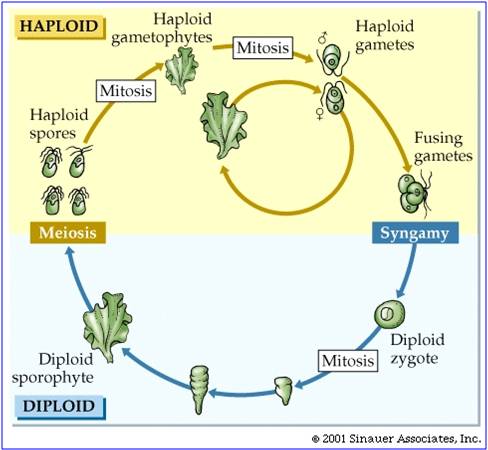 1.
Green Algal Ancestors
1.
Green Algal Ancestors
There is considerable evidence that plants evolved from green algal ancestors. They share the same type of chloroplasts (with chlorophyll a and b), and an intersting life history called "alternation of generation". In this life cycle, a haploid organism (the gametophyte) produces gametes by mitosis. These gametes fuse in a fertilization event, creating a diploid zygote. This diploid zygote divides (typically) mitotically to produce a multicellular diploid offspring. This organism produces haploid spores by meiosis. These spores develop, dividing mitotically to produce the next multicellular haploid generation. So, the ploidy level "alternates" across the generations. In green algae, the sporophyte and gametophyte stages may be "isomorphic" (look the same), like in sea lettuce (Ulva) show at right. In addition, green algae exhibit a range of gamete types, from isogametes that are double flagellated and the same size (as in Ulva) to oogamous species with double-flagellated sperm and immobile large eggs. All plants are oogamous, with large eggs and small sperm. In some gymnosperms and the angiosperms, the sperm have no flagella; in all other plants the sperm have flagella.
2. Colonization of Land: Environmental Differences
 The evolution of plants occurred as photosynthetic organisms acquired traits
that allowed for greater reproductive success on land - out of the ancestral
aquatic environment of algae. So, to understand how plants differ from algae,
and how and why plants evolved various characteristics, you must understand
how aquatic and terrestrial habitats differ. Algae and plants require several
essential nutrients: sunlight, water, and mineral nutrients (like nitrogen,
potassium, magnesium, and phosphorus, to name a few). They also require CO2
and oxygen, as all green algae and plants are photosynthetic aerobes, having
both chloroplasts (for photosynthesis that needs CO2) and mitochondria
(for aerobic respiration that needs O2). Aquatic habitats have ample
water (obviously). Carbon dioxide dissolves in water, so the concentration in
air and water will equilibrate over time. However, CO2 does not diffuse
quickly into or through water, so it may be less abundant locally. Likewise,
oxygen may be in short supply in aqueous habitats, largely because it does not
dissolve as readily. Although oxygen has a concentration of 21% in the atmosphere,
it is often about 7% or less in aquatic habitats, with more near the surface
and less at increasing water depths. In addition, water absorbs light energy
quickly. About 90% of the light energy is absorbed in the first 10m of water;
99% is absorbed by 50-100m. So, photosynthetic organisms must stay near the
top of a water column to get light and oxygen. Unfortunately, the process of
decomposition that recycles mineral nutrients tends to occur at the bottom of
a water column. As such, growth and photosynthesis of aquatic algae is usually
strongly limited by mineral nutrients - either phosphorus, nitrogen, or iron.
(This is why fertilizers running of into water can create huge algal blooms
- the factor previously limiting their growth has been provided, so they grow
like crazy). That is why the open ocean has little photosynthetic activity per
unit area; especially compared to shallower regions like continental margins
and shallow estuaries where upwelling and wave action bring nutrients to the
surface and feed local productivity. So, although aquatic habitats have lots
of water, terrestrial environments provide more sunlight, more oxygen, the same
amount of CO2 (but more equitably distributed and available), and
more mineral nutrients (which are eroding from the rocks or are being recycled
by decomposers and are present in the soil water). And in shallow-water environments
with a supply of nutrients, competition with other algae would become intense,
too.
The evolution of plants occurred as photosynthetic organisms acquired traits
that allowed for greater reproductive success on land - out of the ancestral
aquatic environment of algae. So, to understand how plants differ from algae,
and how and why plants evolved various characteristics, you must understand
how aquatic and terrestrial habitats differ. Algae and plants require several
essential nutrients: sunlight, water, and mineral nutrients (like nitrogen,
potassium, magnesium, and phosphorus, to name a few). They also require CO2
and oxygen, as all green algae and plants are photosynthetic aerobes, having
both chloroplasts (for photosynthesis that needs CO2) and mitochondria
(for aerobic respiration that needs O2). Aquatic habitats have ample
water (obviously). Carbon dioxide dissolves in water, so the concentration in
air and water will equilibrate over time. However, CO2 does not diffuse
quickly into or through water, so it may be less abundant locally. Likewise,
oxygen may be in short supply in aqueous habitats, largely because it does not
dissolve as readily. Although oxygen has a concentration of 21% in the atmosphere,
it is often about 7% or less in aquatic habitats, with more near the surface
and less at increasing water depths. In addition, water absorbs light energy
quickly. About 90% of the light energy is absorbed in the first 10m of water;
99% is absorbed by 50-100m. So, photosynthetic organisms must stay near the
top of a water column to get light and oxygen. Unfortunately, the process of
decomposition that recycles mineral nutrients tends to occur at the bottom of
a water column. As such, growth and photosynthesis of aquatic algae is usually
strongly limited by mineral nutrients - either phosphorus, nitrogen, or iron.
(This is why fertilizers running of into water can create huge algal blooms
- the factor previously limiting their growth has been provided, so they grow
like crazy). That is why the open ocean has little photosynthetic activity per
unit area; especially compared to shallower regions like continental margins
and shallow estuaries where upwelling and wave action bring nutrients to the
surface and feed local productivity. So, although aquatic habitats have lots
of water, terrestrial environments provide more sunlight, more oxygen, the same
amount of CO2 (but more equitably distributed and available), and
more mineral nutrients (which are eroding from the rocks or are being recycled
by decomposers and are present in the soil water). And in shallow-water environments
with a supply of nutrients, competition with other algae would become intense,
too.
In this context, land looks like a much better habitat for photosynthetic organisms than aquatic habitats. We can appreciate why selection might favor organisms that could colonize this environment. However, there is one significant hurdle - terrestrial environments are very very dry. Living tissue is mostly water; in order to survive in a terrestrial environment, living systems had to evolve mechanisms for reducing water loss. There are other problems that a deficit of water creates, too, like for reproduction. Algae and animals have swimming gametes; how can they reproduce on land? As we consider different groups of plants, we will focus on how plants evolved characteristics that solved these problems.
 3.
Plant Evolution: Adapting to Land
3.
Plant Evolution: Adapting to Land
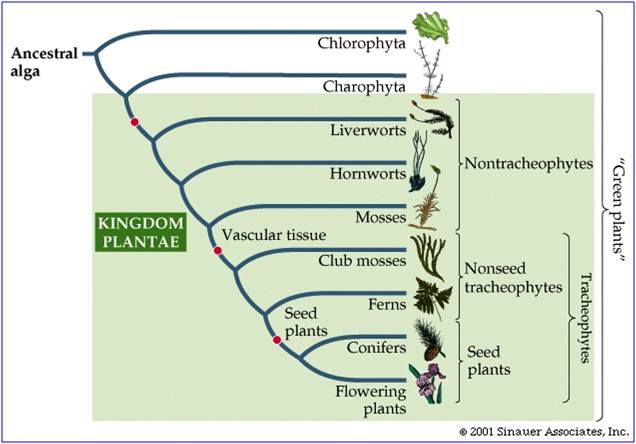
The dominant plants on the planet today are the flowering plants (Angiosperms) and the conifers (a group of Gymnosperms). Both groups are fully terrestrial (except those that have recolonized water!), meaning that no stage of their life cycle requires an aquatic habitat to survive. They have a waxy cuticle on their leaves that reduces water loss, and they also have pores ("stomates") that close during dry conditions to reduce water loss still further. They have vascular tissue for the efficient transport of water from the roots to leaves ("xylem" tissue) and to transport sugars from leaves to the rest of the plant ("phloem" tissue). The xylem has cellulose and lignin in the cell walls, creating rigid tissue that can support the weight of the plant. In addition, these plants have non-motile sperm; the sperm is carried to the egg by the male gametophyte organism, the pollen grain. All of these characteristics are adaptations to life on land: reducing water loss, increasing the efficiency of water transport, and reducing dependancy on water as a necessary medium for gamete transport or bodily support. These adaptations did not evolve at once. Rather, through the evolution of different new plant groups, plants evolved and accumulated these adaptations. Today, although angiosperms dominate the plant kingdom, there are still representatives of more ancient groups. As we move through these groups, we will emphasize the major evolutionary innovations of each group.
4. Primitive Plants - the "non-tracheophytes"
 a.
Groups
a.
Groups
These groups are not a 'monophyletic' clade, but we will consider them together. They are the "liverworts", the "hornworts", and the "mosses". You are certainly aware of mosses, but liverworts are quite common, too, living on streambanks or on rocks in streams. Hornworts are a bit less common, but are also found in moist areas, including the bark of some trees.
b. Characteristics
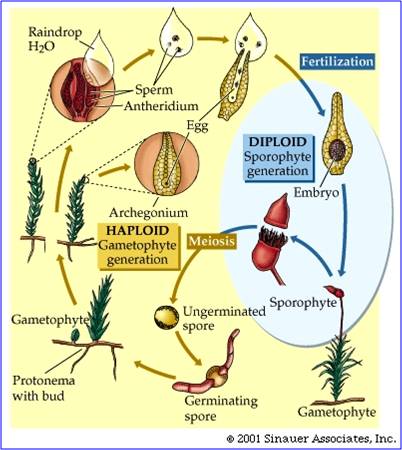
These groups are all non-vascular, meaning that they do not have xylem or phloem. As a consequence, water and sugar transport in the plants is from cell to cell and it is not very efficient. So, the plants must remain small and low to the ground - the source of their water. They also produce swimming sperm, and so must be covered by a thin film of water in order to reproduce. They do not have true leaves or roots (defined as having veins), and so water absorption is inefficient, too. In short, these plants are limited to living in very moist areas, and they must be covered by at least a thin film of water at some time during the year. These plants do have some adaptations for life on land that distinguish them from their algal ancestors, however. They secrete a thin oily cuticle that protects them a bit from water loss. In addition, hornworts and moss sporophytes have stomates - pores in their 'leaflets' to regulate gas exchange and water loss. Their life cycle is dominated by the haploid gametophyte generation; that means that when you are looking at a green moss plant or a liverwort, you are looking at a haploid gametophyte organism. The sporophyte organism is smaller, and is physiologically dependent on the haploid gametophyte parent. In moss, the sporophyte grows out of the archogonium (egg-bearing structure) on the top of the female moss gametophyte. It is brown, and receives the energy it needs to grow from the parent plant. The capsule that it produces is where meiosis occurs, reducing ploidy by 1/2 and producing the haploid spores that will each develop into new haploid gametophyte plants.
c. History
The evolutionary history of these plants is scant, because they do not produce thick, lignified cell walls that fossilize. However, scientists working in Argentina unearthed what they believe to be Liverwort spores that date to approximately 470 mya. This is the earliest fossil record of a terrestrial plant group.
5. Tracheophytes
 a.
Characteristics
a.
Characteristics
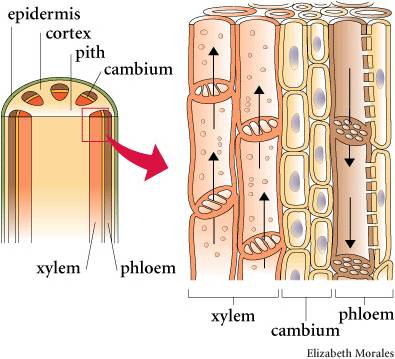
As the name denotes, these are the vascular plants that have true xylem and phloem tissue. Xylem tissue consists of cells that have died, leaving behind their highly lignified cell walls. The ends of these vessel elements have holes, allowing water to pass through like a straw. The evolution of xylem allows for efficient transport of water up a plant. This occurs through root pressure and evapotranspiration from leaves. By actively transporting salts into the xylem cavity, water follows by osmosis from living root tissue and water pressure 'pumps' water up the stem. At the other end of the xylem vessels, water is evaporating off leaves to the environment, creating a negative pressure that 'sucks' water up the stem. Because of the narrowness of the vessel elements, water also moves up by capillary action. All this means that plants could grow taller. This is advantageous for gathering light (the plant is not subject to shading by neighbors) and dispersing spores, but not too great for dispersing swimming sperm.
As such, it is not surprizing that the sporophyte generations becomes the dominant life history stage in tracheophytes. The gametophyte can't take advantage of larger size, because the plant needs to be covered by water for the sperm to swim. The sporophyte, however, benefits from being tall because the increased wind disperses spores farther.
b. Groups:
There are two major groups of tracheophytes, the Lycopodiophytes and the Euphyllophytes. These groups represent true evolutionary clades. They are distinguished by the types of leaves they have. The more primitive group, the Lycopodiophytes, have leaves with one unbranched vein. The Euphyllophytes (which means plants with true leaves), have leaves with many, often branching veins. The Lycopodiophyes include the clubmosses, spikemosses, and quillworts. The Euphyllophytes include the Monilophytes (ferns and kin), and the seed-bearing plants (Gymnosperms and angiosperms).
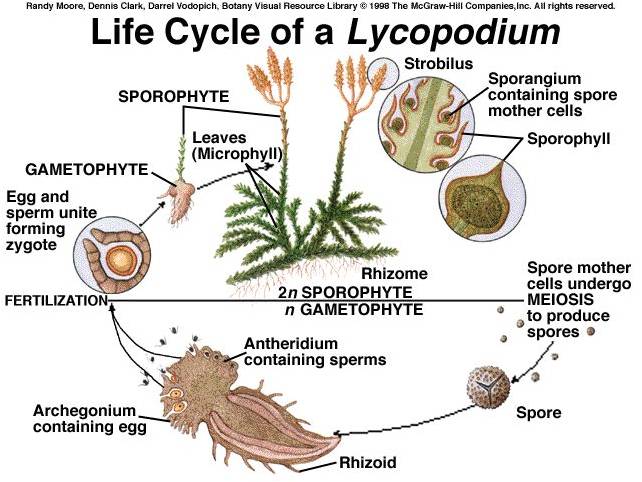 i.
Lycopodiophyta: These are the most primitive living vascular
plants, descended from a very diverse clade that dominated through the late
Silurian, Devonian, and Carboniferous Periods (400-300 mya) of the Paleozoic
Era. The presence of vascular tissue allowed their sporophytes to grow tall;
indeed, the 'arborescent' (tree-like) canopy of Devonian and Carboniferous forests
were clubmosses nearly 100 ft tall. Today, clubmosses are typically under a
foot in height, forming a lush carpet in the herb layer of moist forests. The
life cycle shows this inversion and dominance of the sporophyte. When you are
looking at a green clubmoss plant, you are seeing the diploid sporophyte. It
bears cone-like structures called strobili, and in those structures the sporangia
produce haploid spores by meiosis. The spores germinate, producing a free-living
gametophyte that lives in the soil. Archegonia and antheridia produce eggs and
sperm, respectively, by mitosis. The swimming sperm fertilize the eggs in the
archegonia, and the developing sporophyte grows out of its gametophyte parent,
as in the more primitive true mosses. However, this sporophyte is photosynthetically
active and grows to be a large, physiologically independent organism - it is
not dependent on its gametophyte parent as in true mosses.
i.
Lycopodiophyta: These are the most primitive living vascular
plants, descended from a very diverse clade that dominated through the late
Silurian, Devonian, and Carboniferous Periods (400-300 mya) of the Paleozoic
Era. The presence of vascular tissue allowed their sporophytes to grow tall;
indeed, the 'arborescent' (tree-like) canopy of Devonian and Carboniferous forests
were clubmosses nearly 100 ft tall. Today, clubmosses are typically under a
foot in height, forming a lush carpet in the herb layer of moist forests. The
life cycle shows this inversion and dominance of the sporophyte. When you are
looking at a green clubmoss plant, you are seeing the diploid sporophyte. It
bears cone-like structures called strobili, and in those structures the sporangia
produce haploid spores by meiosis. The spores germinate, producing a free-living
gametophyte that lives in the soil. Archegonia and antheridia produce eggs and
sperm, respectively, by mitosis. The swimming sperm fertilize the eggs in the
archegonia, and the developing sporophyte grows out of its gametophyte parent,
as in the more primitive true mosses. However, this sporophyte is photosynthetically
active and grows to be a large, physiologically independent organism - it is
not dependent on its gametophyte parent as in true mosses.
ii. Monilophytes: These are the ferns and their allies; they form a a monophyletic group of euphyllophyte plants (with true leaves) that, like all plants we have considered, do not produce seeds and have swimming sperm. The other euphyllophytes DO produce seeds, and form the monophyletic group called the Spermatophytes. Some of the Monilophytes, like Equisetum and Psilotum, don't look much like a fern... however, their life cycles, presence of sporangia, and genes show their relationship to the ferns.
Diversity:
The Ophioglassales and Psilotales are very unusual ferns - they have a highly reduced morphology such that Ophioglossum has but a single, 2-part frond and Psilotum lack any fronds (leaves) at all. Psilotum was thought to be related more closely to Lycopods until genetic analyses revealed a closer association with ferns.
 |
 |
| Ophioglossum | Psilotum |
The Equisetales are another group of fern relatives, known as "horsetails" . They also have a reduced morphology that made their taxonomic placement difficult before genetic analyses reveal their close relationship to ferns. The leaves of horsetails are very reduced, like in Psilotales - in fact, the leaves are the small, brown, triangular structures that stick up at each 'node' along the stem. The leaves do not photosynthesize; that function is performed by the stem. There are branching types of horsetails, too, with lateral branches arising as whorls from each node of the stem. Horsetails are sometimes called "scouring rushes"; they are very tough and often have accumulated silica in their tissues. They were used by native Americans and early colonists and settlers to scrub pots clean. Giant horsetails in South America can grown 12 ft tall.
Another odd group are the Salvinales. These are water ferns, and they are actually the sister group to the true ferns. They have diverged morphologically to adapt to aquatic habitats.
 |
 |
 |
| Brown leaves | branches in whorls | Salvinia - a fern relative adapted to aquatic habitats |
The more 'typical' ferns fall into three groups. The Osmundales have dimorphic fronds - one type is broad and green and photosynthetic, the other bears the spores. The cinnamon fern is a classic representative of this group, and it's tall spikes of spore-bearing leaves are easy to recognize. The other two clades of ferns are the most species-rich groups. The Cyatheales include the "tree ferns" that can grow over 20 feet tall. Their rhizome grows vertically as a trunk-like structure, and then a whorl of long fronds radiate. Most tree ferns have tropical distributions, but some prefer the cooler habitats of cloud forests at high elevations, and some reach temperate latitudes - such as the Tasmania tree fern. The vast majority of Monilophytes are in the Polypodiales - this group includes about 1200 of the 1500 species of Monilophytes, and includes the typical ferns that you know, like bracken fern and staghorn fern.
 |
 |
  |
| Osmunda | Tree fern | Bracken fern and staghorn fern - Polypodiales |
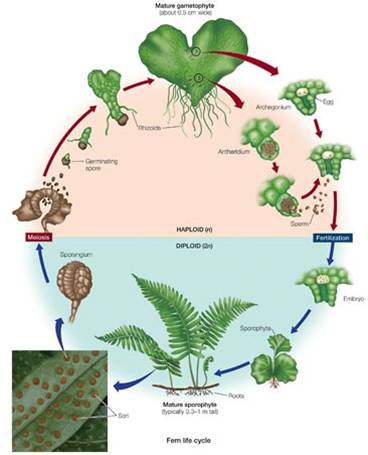 The
fern life cycle is very similar to the clubmosses; indeed, these groups were
sometimes grouped together. Ferns have a dominant sporophyte that produces true
leaves (fronds). In many species, the leaves bear sporangia underneath, which
produce spores by meiosis. The haploid spores are released, and they develope
into small, independent gametophytes that live in the leaf litter. The gametophytes
bear antheridia and archegonia, that produce sperm and egg, respectively, by
mitosis. The flagellated sperm swim to the egg, and the diploid zygote divides
and grows in the archegonium and becomes the large sporophyte.
The
fern life cycle is very similar to the clubmosses; indeed, these groups were
sometimes grouped together. Ferns have a dominant sporophyte that produces true
leaves (fronds). In many species, the leaves bear sporangia underneath, which
produce spores by meiosis. The haploid spores are released, and they develope
into small, independent gametophytes that live in the leaf litter. The gametophytes
bear antheridia and archegonia, that produce sperm and egg, respectively, by
mitosis. The flagellated sperm swim to the egg, and the diploid zygote divides
and grows in the archegonium and becomes the large sporophyte.
The great swamp forests of the Carboniferous (360-286 mya, sometimes split into Mississippian and Pennsylvanian, as in the ppt) were dominated by lycopods like Lepidodendron. Tree ferns were common in the understory, and smaller lycopods and ferns were common in the herb layer, as they are today. As the large plants in these forest died, they fell into the shallow swamps and were covered by sediments. Instead of being decomposed, the organic remains were fossilized, becoming our major coal deposits and giving the period its name (Carboniferous). This huge transfer of carbon to the lithosphere changed the atmospheric balance of carbon dioxide and oxygen. The photosynthetic plants were producing lots of oxygen and were absorbing lots of carbon dioxide, coverting carbon dioxide to glucose and cellulose. Typically, the action of aerobic, decomposers would consume some of this oxygen during their respiration, and would break down the cellulose and glucose into carbon dioxide. Thus, the effects of photosynthetic organisms on the atmosphere is balanced by the effects of decomposers. However, the sequestering of this plant biomass beneath sediment meant that it wasn't metabolized and recycled; it was protected from decomposers. So, oxygen concentrations continued to rise and carbon dioxide levels dropped. Some biologists believe that the high oxygen levels may have been a factor that allowed for the largest terrestrial arthropods to survive - including a dragonfly with a 2 ft wingspan and a millipede 6 feet long. Because arthropods respire by simple diffusion, large arthropods could only receive enough oxygen to maintain their metabolic demands if the concentration gradient was so large that oxygen poured deep into tissues.
In the Permian Period, the continents united into the supercontinent of Pangaea. This land mass had a lower ratio of coastine to land area than multiple independent continents, and so most of the land mass of Pangaea was far from coastal areas. As a consequence, the terrestrial environment went through a drying period; a period which favored new groups of organisms that had evolved characteristics that would give them a selective advantage in dry environments. These organisms were the gymnosperms and the reptiles, and their adaptations were seeds and desiccation-resistant shelled eggs, respectively.
 iii.
Gymnosperms: The
gymnosperms evolved during the Carboniferous but came to dominate terrestrial
landscapes during the Permian Period, when the unification of Pangaea created
a drier terrestrial climate. They would continue to dominate through the first
part of the Mesozoic. Modern gymnosperms include the conifers (pines, fir, spruce,
hemlock, yew, redwoods,cedars, cypress, larch, etc.), the cycads, and the Ginkgo
(one relict species). Another group, the Gnetales, contains some very unusual
gymnosperms that have some characteristics of angiosperms. The gymnosperms probably
evolved from an earlier group of plants called the "Seed ferns". Seeds
ferns were the first plants to produce seeds and pollen; characteristics which
would allow their descendants to dominate terrestrial habitats. The seed ferns
are an extinct group, so we will use the modern Gymnosperms to describe the
importance of these key evolutionary innovations.
iii.
Gymnosperms: The
gymnosperms evolved during the Carboniferous but came to dominate terrestrial
landscapes during the Permian Period, when the unification of Pangaea created
a drier terrestrial climate. They would continue to dominate through the first
part of the Mesozoic. Modern gymnosperms include the conifers (pines, fir, spruce,
hemlock, yew, redwoods,cedars, cypress, larch, etc.), the cycads, and the Ginkgo
(one relict species). Another group, the Gnetales, contains some very unusual
gymnosperms that have some characteristics of angiosperms. The gymnosperms probably
evolved from an earlier group of plants called the "Seed ferns". Seeds
ferns were the first plants to produce seeds and pollen; characteristics which
would allow their descendants to dominate terrestrial habitats. The seed ferns
are an extinct group, so we will use the modern Gymnosperms to describe the
importance of these key evolutionary innovations.
- Characteristics
The seed: The plants that we have examined so far produce the diploid zygote on the female gametophyte - in the vase-like archegonium. As such, the dispersal phase of the life cycle is the spore. Spores are single celled, and although they may be dessication resistant, once they germinate and begin to grow, they must be able to immediately meet their biological needs with material they absorb from the environment. In the "seed plants" (seed ferns, gymnosperms, and angiosperms), the female gametophyte is retained on the sporophyte plant, and becomes the dispersal vessel for its offspring, the next generation of embryonic sporophyte. Here's what happens: The sporophyte produces two types of spores, large and small. Inside the ovule, a 2n cell (megasporocyte) divides by meiosis. The cytoplasm is divided unequally, producing three reduced non-functional cells and one large functional spore - the megaspore. This megaspore, nested within the ovule, divides mitotically and developes into the female gametophyte. The female gametophyte produces the egg, located at one end of the female gametophyte, within the ovule. Fertilization of the egg by sperm forms the diploid zygote. This divides to become the diploid embryo, all still surrounded by haploid gametophyte tissue, within original 2n ovule tissue. This is the seed: 2n ovule tissue (seed coat), surrounding 1n female gametophyte tissue, surrounding the 2n embryo. The gametophyte tissue stores energy in the form of fats and lipids. When the seed is dispersed, the developing embryo now has a source of nutrients to use to produce its initial roots and leaves and become established in the environment. The seed represents a way that the plant can invest MORE ENERGY in each offspring, increasing the probability of survival.
Pollen: Correlated with the production of seeds is the production of pollen. As mentioned above, the sporophyte produces two types of spores, large and small. Each small spore divides and becomes a multicellular haploid male gametophyte - a pollen grain. These pollen grains are released from the sporophyte plant, and are carried by air currents to female cones which house the female gametophyte. The pollen grain then produces sperm. Cycads and ginkgo produce flagellated sperm, but conifers and gnetales produce non-motil sperm. In either case, the sperm is already at the egg. Obviously, the evolution of pollen, that carries sperm through the environment, meant that water was no longer required as a vehicle for reproduction. Plants were now free to colonize uplant areas away from water.
- Diversity:
There are four groups of living gymnosperms. The cycads are the most primitive group, perhaps reminiscent of the seed ferns from which the gymnosperms evolved. They still have flagellated sperm (an ancestral trait), and stiff, tough, frond-like leaves. Some Cycads even bear megaspores on leaves like the ferns do, although the male microspores and pollen are produced in cones. They also have an unbranched stem that is similar to tree ferns. They do produce true seeds, however, and thus are true (though primitive) gymnosperms. The Ginkgo tree is an unusual species - it is the only species in its entire Order (Ginkgoales) - the last species remaining from a previously diverse group. It also has the primitive trait of flagellated sperm. Pollen is produced in cones, but megaspores are produced in fruit-like structures. The third group - Gnetales, - is the sister group to conifers and is a diverse collection of odd plants. The most diverse group of gymnosperms on the planet today is the conifers. This group includes ~600 of the roughly 900 species of gymnosperms (compared to 300,000 species of flowering plants!!). The group includes the largest plants on earth - the tall redwoods and the massive sequioas. It also includes the oldest plants on the planet - some redwoods and bristlecone pines are over 5000 years old. Conifers include pines, fir trees, spruce, larch, yews, cypress, redwoods and sequioas, hemlock, cedar, and juniper. Conifers, like other gymnosperm groups, arose in the Carboniferous and radiated in the Permian when the climate became drier. Conifers, in particular, radiated during the Mesozoic, filling plant niches emptied by the great Permian extinction. Conifers continue to dominate dry environments, today. They dominate dry coastal habitats where sandy soils reduce water availability, they dominate dry mountain slopes, and they dominate cold habitats at high elevations and high latitudes where water is often frozen and unavailable. Indeed, the boreal forest - one of the largest terrestrial biomes on the planet, is overwhelmingly dominated by conifers. So, although their diversity has diminished and although they have been numerically dominated by flowering plants, gymnosperms continue to be very important components of many terrestrial habitats.
 |
 |
 |
 |
| Cycads have large, fern-like leaves | Ginkgo is a single 'living fossil' species | Gnetales are just weird | Conifers still dominate many habitats |
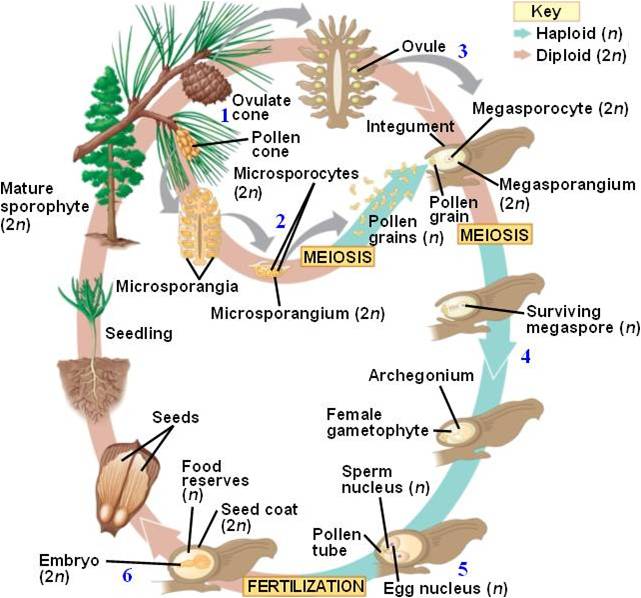 Life
cycle: We will use a pine tree (conifer) as our example of the gymnosperm
life cycle. Consider a mature pine tree - it is the diploid sporophyte generation.
It produces two types of cones, male and female. In the male cones, in compartments
on each "scale", 2n microsporocyte cells go through meiosis and produce
4 equally sized haploid microspores. Each microspore divides by mitosis and
develops into a mature haploid male gametophyte - a pollen grain. Each pollen
grain then produces haploid non-motil sperm by mitosis. Pine trees also have
larger, woody, female cones. In ovules on each scale of these cones, meiosis
occurs and produces a single large haploid spore - the megaspore. The megaspore
divides by mitosis, producing the multicellular haploid female gametophyte.
This megagametophyte produces the egg. There is a pore at one end of the ovule
called the micropyle. When the gametophyte is sexually mature, it secretes a
drop of fluid called the "pollen droplet". Pollen is released from
the male cones and wafts into the female cones. It gets stuck in the fluid droplet,
and as the droplet dries the pollen grain is pulled into the ovule. Inside the
ovule, a tube cell bores through the pollen grain, followed by sperm cells.
Fertilization takes place, forming a diploid zygote within the haploid female
gametophyte tissue. So, there is a diploid embryo, surrounded by haploid gametophyte
tissue, surrounded by a thin covering (seed coat) of grand-parental sporophyte
tissue. This three-layered structure is the seed, and it can be dispersed to
a new environment and gain a footing in that habitat by using the energy packet
along in the female gametophyte tissue.
Life
cycle: We will use a pine tree (conifer) as our example of the gymnosperm
life cycle. Consider a mature pine tree - it is the diploid sporophyte generation.
It produces two types of cones, male and female. In the male cones, in compartments
on each "scale", 2n microsporocyte cells go through meiosis and produce
4 equally sized haploid microspores. Each microspore divides by mitosis and
develops into a mature haploid male gametophyte - a pollen grain. Each pollen
grain then produces haploid non-motil sperm by mitosis. Pine trees also have
larger, woody, female cones. In ovules on each scale of these cones, meiosis
occurs and produces a single large haploid spore - the megaspore. The megaspore
divides by mitosis, producing the multicellular haploid female gametophyte.
This megagametophyte produces the egg. There is a pore at one end of the ovule
called the micropyle. When the gametophyte is sexually mature, it secretes a
drop of fluid called the "pollen droplet". Pollen is released from
the male cones and wafts into the female cones. It gets stuck in the fluid droplet,
and as the droplet dries the pollen grain is pulled into the ovule. Inside the
ovule, a tube cell bores through the pollen grain, followed by sperm cells.
Fertilization takes place, forming a diploid zygote within the haploid female
gametophyte tissue. So, there is a diploid embryo, surrounded by haploid gametophyte
tissue, surrounded by a thin covering (seed coat) of grand-parental sporophyte
tissue. This three-layered structure is the seed, and it can be dispersed to
a new environment and gain a footing in that habitat by using the energy packet
along in the female gametophyte tissue.
With seed fern and conifer evolution, plants had finally become independent of an aquatic environment. No life stage needed liquid water as an environment, and these plant species could colonize the drier areas that were increasingly abundant as the Permian and Mesozoic progressed.
iv. Angiosperms: The angiosperms are the flowering plants; they are the most diverse group of plants on the planet today, represented by approximately 300,000 species and comprising 90% of all plant species. Angiosperms evolved early in the mesozoic, but radiated in the early Cretaceous Period - the last period of the Mesozoic Era. Although their diversity declined as a consequence of the meteor impact 65 mya (known as the K-T boundary because it is used to separate the Cretaceous ("K" - yeah, I know... but Cambrian got "C") of the Mesozoic from the Tertiary ("T") of the Cenozoic Era), they rebounded and dominated most terrestrial habitats. Like their sister group the gymnosperms, they have seeds and pollen. However, they also have two additional distinguishing derived characteristics - flowers and fruits.
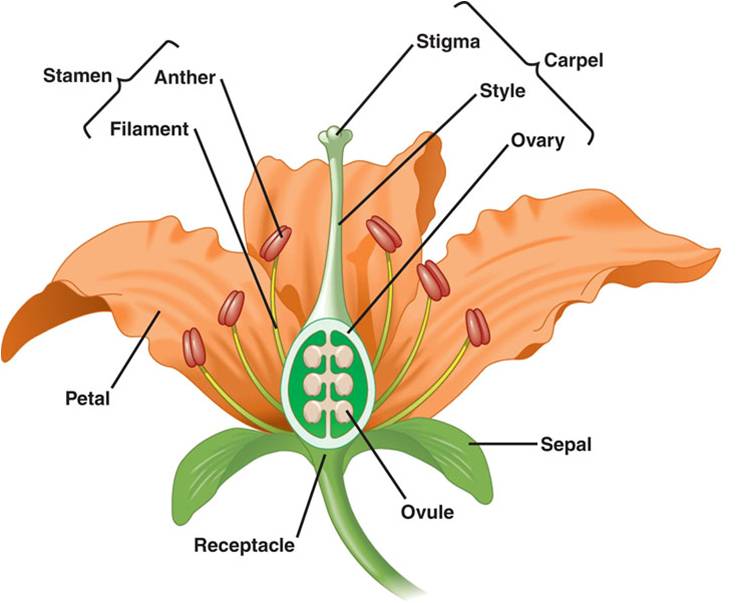 Flowers: A flower is a stem-like structure that bears whorls of leaf-like appendages.
The lowest whorl on this reproductive "twig" is the calyx, composed
of leaf-like sepals that form a protective shealth surrounding the flower bud.
The second whorl of leaf-like appendages is the corolla, containing the leaf-like
petals. The third whorl contains the stamens. Each stamen has a filament that
bears an anther at the end. The anther is where meiosis occurs and haploid microspores
develop into pollen grains. At the center of the flower is the pistil, or carpel.
This is the female portion of the flower, consisting of the stigma (a sticky
surface that receives pollen), the style (a stalk connecting the stigma to the
ovary) and the ovary. The ovary has compartments that contain the ovules. The
structure of flowers can vary tremendously, and may lack carpels, petals, pistils,
or stamens. The number of the parts may vary, also, and 'composite' flowers
may contain thousands of flowers growing together as one big inflorescence -
with some flowers only bearing petals, and other flowers only bearing stamens
or ovaries. A daisy is a good example of a composite - the petals are actually
the product of single 'ray' flowers that each ONLY bear one petal. The yellow
center of a daisy is composed of hundreds of individual 'disk' flowers that
don't have petals but do have stamens and pistils.
Flowers: A flower is a stem-like structure that bears whorls of leaf-like appendages.
The lowest whorl on this reproductive "twig" is the calyx, composed
of leaf-like sepals that form a protective shealth surrounding the flower bud.
The second whorl of leaf-like appendages is the corolla, containing the leaf-like
petals. The third whorl contains the stamens. Each stamen has a filament that
bears an anther at the end. The anther is where meiosis occurs and haploid microspores
develop into pollen grains. At the center of the flower is the pistil, or carpel.
This is the female portion of the flower, consisting of the stigma (a sticky
surface that receives pollen), the style (a stalk connecting the stigma to the
ovary) and the ovary. The ovary has compartments that contain the ovules. The
structure of flowers can vary tremendously, and may lack carpels, petals, pistils,
or stamens. The number of the parts may vary, also, and 'composite' flowers
may contain thousands of flowers growing together as one big inflorescence -
with some flowers only bearing petals, and other flowers only bearing stamens
or ovaries. A daisy is a good example of a composite - the petals are actually
the product of single 'ray' flowers that each ONLY bear one petal. The yellow
center of a daisy is composed of hundreds of individual 'disk' flowers that
don't have petals but do have stamens and pistils.
Flowers evolved to exploit animals for pollen transfer. Wind is capricious - the direction changes, the intensity changes, and even on a windy day only a small fraction of the pollen produced in a male cone will be lucky enough, just by chance, to travel to a female cone. For this reason, gymnosperms typically produce alot of pollen. By contrast, animals move through the environment non-randomly in search of food, shelter, and mates; and all animals can learn. So, by associating pollen with attractants (nectar, fragrance, and colorful floral displays) plants teach animals to visit flowers and increase the frequency of effective pollen transfer. Animals visiting an attractive flower are rewarded with food and quickly learn the association; they then travel in "bee-lines" from one flower to another seeking food, transfering pollen non-randomly and efficiently. This would be particularly useful for plants in the understory of a forest, where wind is minimal. The first angiosperms may have been shrubby, understory species. Animal pollination would also be very adaptive in tropical rainforests or deserts, where members of a single plant species may be very far away (decreasing the chances that a random procees like wind pollination would effectively find a 'mate').
As some angiosperms evolved into taller forms where wind could be used effectively, some were selected to use wind as a pollen-dispersal mechanism - with correlated reductions in flower size. So, wind-pollinated flowering trees (oaks, maples, etc.) and wind-pollinated field plants (open fields are windy environments, too) like grasses often have very small flowers with no colorful petals or sepals. Because they are not attracting animals for pollen dispersal, selection favors organisms that do not invest alot of energy in these structures.
 Plants
can increase the efficiency of pollen transfer even further by limiting flower
visitation to one type of pollinator. A specialized pollinator that visits just
one type of flower is more likely to transfer pollen between plants of the same
species. Likewise, flowers visited by just one type of pollinator are likely
to have nectar present when that pollintor visits. So, "pollination syndromes"
have evolved, representing flower traits that are associated with a particular
type of pollinator. For example, many flowers are red and tubelike. These flowers
are often pollinated by hummingbirds, which see into the red end of the spectrum
better than most insects and have a long beak that can access the nectar at
the base of the flower. Butterfly flowers are often composites (like daisies,
and zinnias), or are clustered in groups (butterfly bush). This allows the butterfly
to land on a cluster and probe many individual flowers at a time with their
long proboscis. Moth and bat pollinated flowers open at night. They are usually
white and very fragrant, attracting their pollinators with scent. Some plants
like skunk cabbage produce flowers that smell like rotting flesh. This attracts
flies that act as pollinators. Some of the most elaborate contrivances by which
plants limit pollination to a single species of pollinator are seen in the orchid
family. Some orchids produce a scent that mimics the sexual pheromones of females
of specific species of solitary wasps. When males perceive the scent, they fly
up the concentration gradient to the flower. In some species, the flower also
mimics the shape and coloration of the female wasp! The deception is so good
that the male attempts to mate with the flower. It gets dusted with pollen,
and flies off. While this may be frustrating for the wasp (who knows), it can't
NOT fly down the next pheromone trail it encounters or it won't mate. So, if
it flies down another trail and is tricked by another plant, pollen is transferred.
The bucket orchid is another strange plant. Two petals have fused into a bucket
that collect water. Pollinators fall in the bucket, and the only exit is through
a tight squeezeway where they are dusted with pollen.
Plants
can increase the efficiency of pollen transfer even further by limiting flower
visitation to one type of pollinator. A specialized pollinator that visits just
one type of flower is more likely to transfer pollen between plants of the same
species. Likewise, flowers visited by just one type of pollinator are likely
to have nectar present when that pollintor visits. So, "pollination syndromes"
have evolved, representing flower traits that are associated with a particular
type of pollinator. For example, many flowers are red and tubelike. These flowers
are often pollinated by hummingbirds, which see into the red end of the spectrum
better than most insects and have a long beak that can access the nectar at
the base of the flower. Butterfly flowers are often composites (like daisies,
and zinnias), or are clustered in groups (butterfly bush). This allows the butterfly
to land on a cluster and probe many individual flowers at a time with their
long proboscis. Moth and bat pollinated flowers open at night. They are usually
white and very fragrant, attracting their pollinators with scent. Some plants
like skunk cabbage produce flowers that smell like rotting flesh. This attracts
flies that act as pollinators. Some of the most elaborate contrivances by which
plants limit pollination to a single species of pollinator are seen in the orchid
family. Some orchids produce a scent that mimics the sexual pheromones of females
of specific species of solitary wasps. When males perceive the scent, they fly
up the concentration gradient to the flower. In some species, the flower also
mimics the shape and coloration of the female wasp! The deception is so good
that the male attempts to mate with the flower. It gets dusted with pollen,
and flies off. While this may be frustrating for the wasp (who knows), it can't
NOT fly down the next pheromone trail it encounters or it won't mate. So, if
it flies down another trail and is tricked by another plant, pollen is transferred.
The bucket orchid is another strange plant. Two petals have fused into a bucket
that collect water. Pollinators fall in the bucket, and the only exit is through
a tight squeezeway where they are dusted with pollen.
Fruits:Many angiosperms also evolved mechanisms to use animals to disperse their seeds. The ancestral condition within the group is probably the production of fleshy, ovary tissue that surrounds the carpels. Like nectar, this fleshy fruit tissue serves as a bribe, encouraging animals to eat the fruits and thereby disperse the seeds. And since animals tend to show a preference for a given habitat type, the seeds get transferred to another place with similar environmental conditions (rather than being strewn haphazardly across the environment by wind). Other fruits are dry - like acorns, grains, burrs, and some beans. If these are small, like in grasses, or have wings like maple samaras, or have feathery structures like milkweed, then the fruit can be dispersed by the wind. Some, like cocklebur, burdock, and goatshead, evolved hooks on their seed pods that could tangle in mammal fur. Some fruits are pea-pod like, and when the pod dries, the seam bursts open and the seeds are "shot out". This is called 'ballistic' dispersal. So, regardless of whether the fruit is animal or wind dispersed, plants have evolve many different ways to get their seeds dispersed and away from the competitive environment beneath the parent plant.
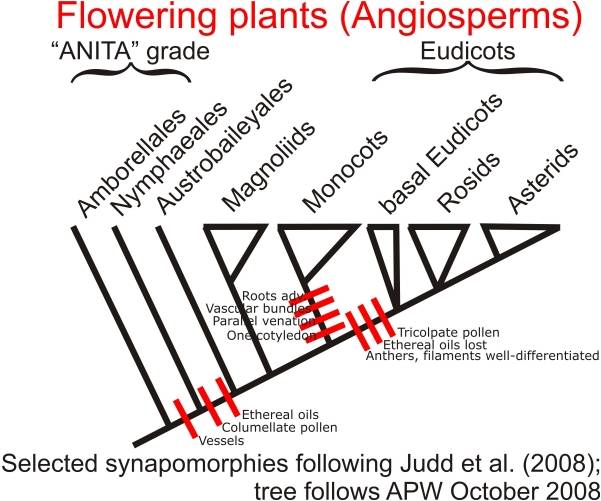 Diversity: The 300,000 angiosperm species have radiated into such a wide variety of shapes
and behaviors that the evolutionary relationships among major groups (which
each contain lots of diversity) is often difficult. However, several groups
do fall out, especially with molecular analyses. Two ancient, basal groups are
the Amborellales ( a small group of a few very primitive species) and the Nymphaeales
(water lilies and their kin). Another ancient group is the Magnoliales, including
the Magnolias and the laurels. Two other large and diverse groups are the monocots
(including grasses, palms, lilies, and orchids), and the dicots (including 70%
of all plants, from roses to oaks, from buttercups to maples). Monocots have
one embryonic leaf (cotyledon), while dicots have two. Also, monocots have leaves
with parallel veins, while dicots usually have leaves with a network of radiating
veins. Monocots typically have flower parts in multiples of 3's, while dicots
have flower parts in multiples of 4's or 5's.
Diversity: The 300,000 angiosperm species have radiated into such a wide variety of shapes
and behaviors that the evolutionary relationships among major groups (which
each contain lots of diversity) is often difficult. However, several groups
do fall out, especially with molecular analyses. Two ancient, basal groups are
the Amborellales ( a small group of a few very primitive species) and the Nymphaeales
(water lilies and their kin). Another ancient group is the Magnoliales, including
the Magnolias and the laurels. Two other large and diverse groups are the monocots
(including grasses, palms, lilies, and orchids), and the dicots (including 70%
of all plants, from roses to oaks, from buttercups to maples). Monocots have
one embryonic leaf (cotyledon), while dicots have two. Also, monocots have leaves
with parallel veins, while dicots usually have leaves with a network of radiating
veins. Monocots typically have flower parts in multiples of 3's, while dicots
have flower parts in multiples of 4's or 5's.
Life Cycle: The generalized life cycle of angiosperms is similar to gymnosperms. The diploid sporophyte produces two types of spores. In angiosperms, however, these can be born by the same structure (perfect hermaphroditic flowers), or different flowers on the same plant (monoecious) or on different plants (dioecious). In the anther, 2n microsporocytes divide by meiosis and produce 4 microspores. Each microspore divides by mitoisis and developes into a pollen grain. Pollen is released from the anther - often on to the body of an animal pollinator visiting the flower. In the ovule, a diploid megasporocyte divides unequally, producing 3 nonfunctional cells and one functional haploid megasporocyte. The nucleus in this cell divides by mitosis through three cycles, producing 8 nuclei. Three aggregate near each pole, and two agregate in the middle of the ovule. The central cell at one end becomes the egg.
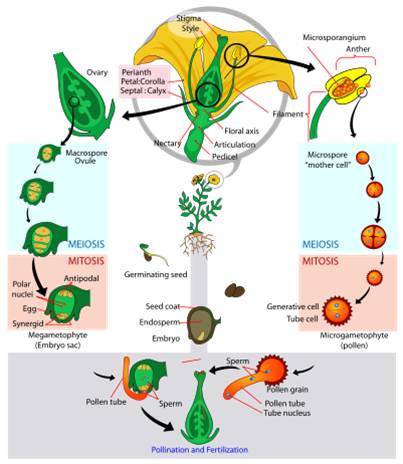 Pollen
lands on the stigma, and one tube cell secretes enzymes that dig a "pollen
tube" through the style to the ovule. It is followed down the tube by two
sperm cells. When they reach the opening of the ovule, one sperm cell fertilizes
the egg and forms the diploid zygote. The other sperm cell fuses with the TWO
polar nuclei in the middle of the ovule, forming a triploid nucleus. This "double
fertilization" is unique to the angiosperms. The triploid nucleus divides,
forming triploid 'endosperm' tissue within the ovule that surrounds the developing
diploid enbryo and provides nutrition. So again, the seed contains three layers
- the ovule of the original diploid sporophyte, the triploid endosperm, and
the diploid embryo.
Pollen
lands on the stigma, and one tube cell secretes enzymes that dig a "pollen
tube" through the style to the ovule. It is followed down the tube by two
sperm cells. When they reach the opening of the ovule, one sperm cell fertilizes
the egg and forms the diploid zygote. The other sperm cell fuses with the TWO
polar nuclei in the middle of the ovule, forming a triploid nucleus. This "double
fertilization" is unique to the angiosperms. The triploid nucleus divides,
forming triploid 'endosperm' tissue within the ovule that surrounds the developing
diploid enbryo and provides nutrition. So again, the seed contains three layers
- the ovule of the original diploid sporophyte, the triploid endosperm, and
the diploid embryo.
6. Summary:
Over the course of plant evolution, organisms evolve characteristics that provided a selective advantage in progessively more desiccating environments, allowing plants to colonize new habitats unoccupied by other plants. The non-tracehophytes evolved a cuticle, and could colonize land and harvest more sunlight than their algal ancestors. Club mosses evolved vascular tissue and a dominant sporophyte, allowing them to grow tall and gain a competitive advantage over smaller non-tracheophytes. Ferns evolved more complex leaves, improving the transport of water and sugars into and out of leaves. The evolution of seeds, the reduction of the the gametophyte, and the elimination of swimming sperm allowed seed ferns and their descendants - the gymnosperms and the angiosperms - to colonize upland areas away from water. Conifers still dominate in the boreal forests of high latitudes; however, the angiosperms dominate in almost all other terrestrial habitats. Their innovations of flowers and fruits exploit animals in the environment, and improve the efficiency of pollen transport and seed dispersal. The evolution of plants is an interesting story of adapting to life on land, to take advantage of the abundant sunlight, oxygen, and mineral nutrients found in this habitat. The are an extraordinarily important, interesting, diverse, and beautiful group of living organisms.
Study Questions:
1. What challenges face an aquatic algae as it tries to acquire light, gases, and nutrients?
2. To a photosynthetic organism, what are the benefits of colonizing land? What is the major "down side"?
3. Why are mosses short? Give two reasons.
4. In clubmosses, the sporophyte is tall but the gametophyte is small. Explain why in terms of the reproductive cells they produce.
5. Ferns and lycopods (clubmosses) dominated during the Carboniferous. Explain how their presence, and their fossilization, changed the atmospheric balance of CO2 and oxygen.
6. Draw the life cycle of a fern.
7. The two major innovations represented in the gymnosperms are seeds and pollen. Explain what each of these structures IS (in the contex of a plant life cycle), and explain how each structure allowed plants to colonize dry habitats.
8. How do showy flowers increase the efficiency of pollen transfer?
9. Describe three pollination syndromes.
10. Where doe the angiosperms that use wind dispersal live? Why have they evolved small flowers?
11. What is the adaptive advantage of fleshy fruit production?