 Overview:
Overview:
Overview:
As curious scientists, we are always interested in beginnings - what causes this or that. Events that occur repeatedly are obviously easier to study through science than events that occur once. In fact, for events that occur once, all we can really do in science is establish correlation - we can't replicate the event, so we can't test it directly. But, we can ask post-dictive hypotheses and make other tests of important explanatory elements. For instance, general relativity and the big bang theory predicted more than just "the expansion of the universe". It predicts the existence of black holes and the behavior of atoms and heavenly bodies under new situations. These have been tested and verified, so we have confidence in the explanatory power of general relativity, in general (ha-ha). This gives us greater confidence in it's explanatory relevance to the universe, as well, even though we can't "create another universe" and test it completely. The origin of life, like the origin of the universe, is such a question. As far as we know, life originated once - on a rocky planet orbiting a sun in a distant arm of a spiral galaxy.
Before we get started presenting ideas about how this process may have occurred, it is important to distinguish "origin of life" hypotheses from "evolution". These are two very different issues with different degrees of empirical support. Evolution is "a change in the genetic structure of a population". Evolution explains the way life changes through time, and the creation of new species as a result of divergence from existing species. AS SUCH, evolution assumes the existence of life. Evolutionary theory, explicitly, is independent of hypotheses regarding how life began. However, evolution does suggest that life is related by common ancestry to some initally successful type of life. Again, however, how those initial life forms arose is a different question. It is analogous to the Copernican Theory of a sun-centered universe. That theory describes how the planets move, but makes no claim to explain how the planets formed. Nonetheless, long before anyone had a good idea about how the solar system formed, the Copernican Theory was accepted as a useful explanation of the movement of the planets. Likewise, evolution describes how life changes over time, and is not really dependent on an particular explanation of how life first arose. And, like the Copernican Theory, Evolutionary Theory has been accepted as a useful description of how life changes through time without yet knowing how life first arose. Critics of evolution often confuse these points, suggesting that evolution is suspect because we do not know how the first life arose. Hopefully, after the last couple weeks, you can see the difference between "evolutionary theory" and "origin of life hypotheses".
The origin of life presents some very interesting (if not novel) problems. For example, even the simplest living cells are very very complex, and are composed of interdependent subsystems. It is difficult to postulate how this complex system could have arisen by stepwise, evolutionary processes from non-living systems. Indeed, having just learned about protein synthesis, one of the more interesting problems may occur to you: if DNA codes for proteins, but protein enzymes are needed to replicate and transcribe DNA, how could one come before the other? It would seem that both proteins and DNA would have to evolve at the same time for the system to work. You would be correct to conclude that this would be highly unlikely. However, our inability to see an answer does not mean that an answer does not exist. Indeed, hard-working scientists have actually figured out a very reasonable hypothesis for how our current genetic system could have evolved - and we will see that hypothesis in this lecture.
A. The Early Atmosphere
 When the Earth formed, any
free hydrogen or helium gas would have been too light to have been held by the
Earth's gravitational field at the time, and was lost to space. The gases released
by volcanoes would have been held as the first permanent atmosphere. Because
the process of vulcanization and the burning of rock is thought to be the same
today as it was throughout Earth history, we can determine the composition of
volcanic gases today and have a pretty good idea of the composition of the early
atmosphere. These volcanic gases are carbon dioxide (CO2), and water
vapor (H2O), Hydrogen gas (H2), Nitrogen gas (N2),
Hydrogen sulphide (H2S), and trace amounts of methane (CH4)
ammonia (NH3), and HCl.
By about 4.4 by, geologists surmise that the earth may have cooled to the point
that water vapor would condense; raining down to fill the ocean basins. However,
through the period of heavy bombardment by meteorites, it is likely that much
of Earth's surface waters were repeatedly vaporized by the energy of these impacts.
After the period of bombardment, the ocean basins filled for good. Liquid water
has had a profound effect on the chemistry of our planet. For example, many
of the volatiles released by volcanoes went into solution easily - such as CO2,
HCl, and SO2. But N2 is not soluble so it remained behind
and dominates our atmosphere (~78%). The sulphuric, nitric, and carbonic acids
that precipitate with the condensing water dissolve rock, washing the dissolved
salts to the ocean where their concentration accumulates.
When the Earth formed, any
free hydrogen or helium gas would have been too light to have been held by the
Earth's gravitational field at the time, and was lost to space. The gases released
by volcanoes would have been held as the first permanent atmosphere. Because
the process of vulcanization and the burning of rock is thought to be the same
today as it was throughout Earth history, we can determine the composition of
volcanic gases today and have a pretty good idea of the composition of the early
atmosphere. These volcanic gases are carbon dioxide (CO2), and water
vapor (H2O), Hydrogen gas (H2), Nitrogen gas (N2),
Hydrogen sulphide (H2S), and trace amounts of methane (CH4)
ammonia (NH3), and HCl.
By about 4.4 by, geologists surmise that the earth may have cooled to the point
that water vapor would condense; raining down to fill the ocean basins. However,
through the period of heavy bombardment by meteorites, it is likely that much
of Earth's surface waters were repeatedly vaporized by the energy of these impacts.
After the period of bombardment, the ocean basins filled for good. Liquid water
has had a profound effect on the chemistry of our planet. For example, many
of the volatiles released by volcanoes went into solution easily - such as CO2,
HCl, and SO2. But N2 is not soluble so it remained behind
and dominates our atmosphere (~78%). The sulphuric, nitric, and carbonic acids
that precipitate with the condensing water dissolve rock, washing the dissolved
salts to the ocean where their concentration accumulates.
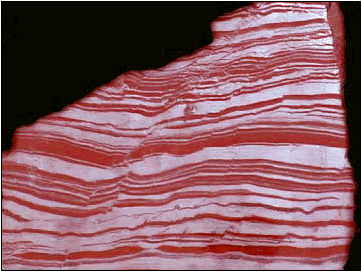
One of the most interesting characteristics of the early atmosphere is that it contained only trace amounts of oxygen gas (O2). How do we know? Well, first, there is very little O2 in volcanic gases - the probable source of the earth's first atmosphere. Second, sedimentary rocks older than about 2 by do not have oxidized minerals. Volcanic rock contains lots of iron. As these iron atoms are eroded and placed into solution, they dissolve and stay in solution as long as oxygen is absent, accumulating in the oceans. When oxygen is present, the iron atoms react with oxygen to form iron oxides, which are not soluble in water and precipitate out. The first deposits of iron-rich sediments date to about 2.0-2.5 by, dating the time when oxygen gas - produced by newly evolved 'oxygenic' photosynthetic pathways - was first released into the oceans and atmosphere by photosynthetic bacteria. Indeed, there was so MUCH dissolved iron in the oceans that it reacted with most of the oxygen released by photosynthetic organisms; soaking the oxygen up and precipitating it out of solution, too. Most of the oxygen in the earth system is bound in these sedimentary deposits. It wasn't until about 1.0 by ago that the dissolved iron had precipitated and the free oxygen gas could diffuse from solution and begin to accumulate in the atmosphere. Even then atmospheric concentrations remained below 5% until about 650 mya.
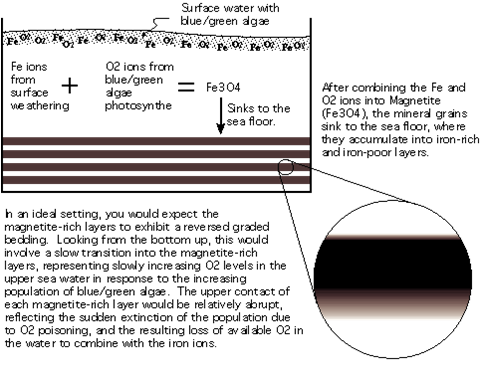
The absence of oxygen gas meant that the early atmosphere was a reducing chemical environment. Although there has been some debate over this claim (with some scientists suggesting that the atmosphere was, at best, only weakly reducing), more recent work supports the claim of a reducing atmosphere. This is important, chemically, because it means that complex biological molecules could accumulate - without being destroyed by oxidative processes.
B. The Formation of Biologically Important Molecules
Life is complex, and is composed of complex subunits. Reductionism and probability imply that complex cells probably did not form spontaneously with all of their structures and functions intact. Rather, naturalistic, reductionistic models hypothesize that subunits would have to form first and then come together. At the most fundamental level, we might expect monomers to form, and then polymers, and then cell-like structures.
1. The source of biologically important monomers
 a.
Spontaneous generation on Earth
a.
Spontaneous generation on Earth
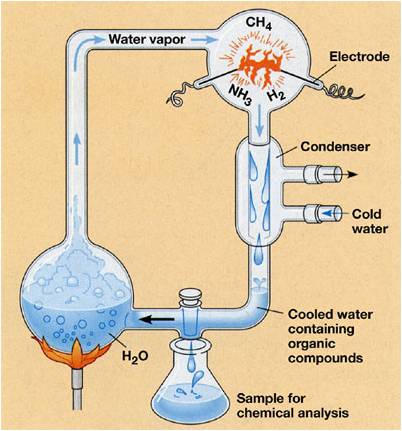
In the 1920's, Alexander Oparin and J. B. S. Haldane independently hypothesized that biologically important monomers like amino acids and sugars might form spontaneously if a mixture of volcanic gases were bombarded with energy in a reducing environment. Curiously, this hypothesis was not effectively tested until 1953, when Stanley Miller and Harold Urey sent electric charges through gaeous mixtures of H2O, CH4, NH3, CO, and H2. In their closed system, the gases were condensed and dissolved solutes accumulated in solution. After a week of running the experiment, they found several amino acids in solution; confirming the Oparin and Haldane hypothesis that biologically important monomers could form spontaneously. Although scientists challenged the appropriateness of their gas mixture (volcanic gases contain other gases, too) and the constantly high application of electric discharge, more recent experiments using a wide variety of volcanic gases and other sources of energy also produced a wide variety of amino acids and all other biologically important molecules including nitrogenous bases, ribose, deoxyribose, fatty acids, phospholipids, and a variety of other sugars and other organic molecules. Although Miller and Urey surmised that the initial source of energy might be lightening strikes at the surface of the earth, other scientists have considered other locations for the synthesis of biologically important molecules. Günter Wächtershäuser (1990) suggested that hydrothermal vents might be a likely spot, where rich mixtures of volcanic gases are released under high heat and pressure directly to the surrounding water. Critics suggested that temperatures may be too hot for molecules to persist, but it is possible that cooler hydrothermal vents called "white smokers" might have been involved.
b. Seeding from Space
In 1969, the Murchison Meteorite fell to earth in Australia. Examinations revealed the presence of amino acids and other organic molecules, leading some scientists to suggest that the period of heavy meteorite bombardment could have seeded the earth with biologically important molecules necessary for life to emerge.
 2.
Formation of Polymers
2.
Formation of Polymers
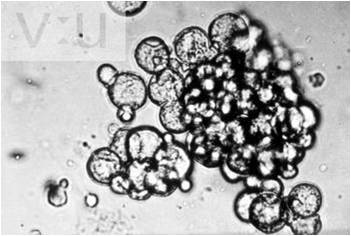
Polymerization of all biologically important molecules works by the simple proces of dehydration synthesis, and it didn't take long for scientists to polymerize amino acids spontaneously. In the 1960's, shortly after the Miller-Urey experiments, Sidney W. Fox discovered that amino acids would polymerize into small spheres composed of proteins - 'proteinoids microspheres' at fairly low temperatures (70oC) if phosphoric acid was present as a catalyst. He found similar microspheres in volcanic lava on Hawaii, and suggested that this might be a spontaneous process by which proteins could have formed. These microspheres are differentially permeable, and they also bud (split into two spheres) when they enlarge. Fox thought these could have been the first 'membranes', creating a unique chemical environment in which other reactions could occur. In the 1960's Cairns-Smith demonstrated that clay particles could act as a polymerization template. The specific charge characteristics on a clay surface would bind a non-random (but not explicitly sequence specific) series of amino acids, and drying at warm temps would cause polymerization. In 2004, Szostak showed that RNA nucleotides coul polymerize on clay surfaces, as well. Again, proponents of a hydrothermal vent origin of life argue that these types of polymerization reactions would be quite possible (and more stable) under the conditions at these vents.
C. Acquiring the Characteristics of Life
A soup of biologically important molecules is not a living cell. Once again, we are confronted by a seemingly impossible process of creating complexity from simple parts. As daunting as this questions sounds, scientists have made considerable progress in shedding light on how this process might have happened. In short, there are three basic functions that a cell must perform: 1) it must be be a bounded system that, as a consequence of differential permeability, can become different from its environment; 2) it must acquire matter/energy and harvest energy through metabolic process; and 3) it must have a genetic system that can co-ordinate this metabolic activity and also replicate. Surprizingly, there are very good hypotheses about how each of these steps might have occurred. However, there is still little agreement on the order in which these steps would have occurred. So, we will present these steps separately, but no order is implied.
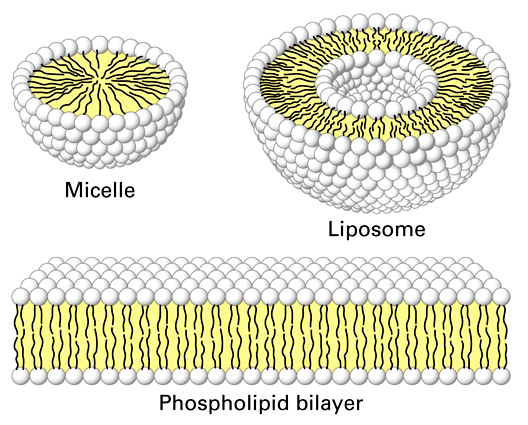 1.
Evolution of a membrane
1.
Evolution of a membrane
When Hydrogen cyanide and formaldehyde are added to Miller Urey experiments, phospholipids form spontaneously. In a turbid environment, wave action will cause these films to ball-up as single-layer micelles and as phospholipid bilayers. As you know, these bilayers form the semi-permeable membrane found in all living cells. So, the formation of a membrane is the easiest and most well-understood piece of the puzzle.
2. Metabolic Pathways
a. Reverse Evolution
Metabolic pathways are often complex; a useless substrate is modified and changed into something else by an enzyme (enzyme 1). This intermediate is modified by another enzyme (enzyme 2), and is converted into another intermediate. Finally, this intermediate is modified by a third enzyme (enzyme 3), and a useful functional product is made.
Substrate A ---> intermediate B ---> intermediate C ---> useful product D
|
|
|
enzyme 1 used enzyme 2 used
enzyme 3 used
- It seems like all three enzymes have to be present at once for the useful product to be made. It couldn't occur by a stepwise process, because a system with just the first step (substrate A to intermediate B) isn't functional - there is no useful product and it dies! So, it seems that all the steps have to be present at once. Again, this is the argument of intelligent design. Do you recognize it? A stepwise process is impossible; this is too complex; everything has to be present together for it to work - be it the eye, a new species, a new gene, or now, metabolic pathways. It is all Paley's old argument, wrapped in new clothes. Well, as with the evolution of the eye, there IS another possibility.
Suppose useful product D is present in the primordial environment. All the protocells are gobbling it up and using it. Well, we might expect the concentration of D to drop. If the useful product "D" is now in short supply, then selection will favor protocells that can make it out of something else - making it on their own and not relying on environmental supply. Maybe they can absorb something similar, like substance C, and "tweak" it chemically into D. So, selection will favor the evolution of the pathway: C--> D, even if C can't be used for anything else.
So, if this pathway is adaptive, then cells with this pathway increase in frequency. Generations later, only protocells with this pathway exist, and all the cells are competing for substrates D (which they can still use when they find it) AND substrate C, which all cells convert to useful product D.
Now, as substrate C decreases in supply (because cells are using it to make D), selection will favor protocells that can convert B into C, and thus into D: B--> C --> D.
Thus, useless intermediate substrates are added to the process in a sequence that is the "reverse" of the direction of the pathway, itself; and a complex pathway of A--> B --> C --> D evolves by a stepwise process.
- Many pathways are fundamentally quite similar - electron transport chains, etc. As is often the case in biological systems, once a functioning system evolves it is often "co-opted" for other functions - like electron transport systems are used in photosynthesis and respiration.
a. Hydrogen Pumps and Reactions
The problem with most Miller-Urey experiments is that the source of energy is episodic - a blast of energy that causes reactions and then stops. But in life, chemical reactions are continuous, and require a continuous source of energy and reactants. The "white smoker" hyothesis provides these conditions. Hot, alkali (basic) water is percolating up through the smoker. On the other side of the very thin inorganic film ('membrane') is cooler acidic water. Electrons are stripped from H2 by the metal-sulphur complexes in the mineral membrane, and are passed to the H+ ions in the acidic water - which reacts with CO2 dissolved in the water to create organic molecules. They are flushed to cavities where they concentrate and can interact. In addition, the H+ ions outside can flood across the membrane to the basic solution inside: a H+ concentration gradient that can be exploited for chemical reactions and synthesis of ATP. And since the vent is always pumping more alkali solution up, the gradient is constant - a constant supply of energy and a constant synthesis of organic molecules.
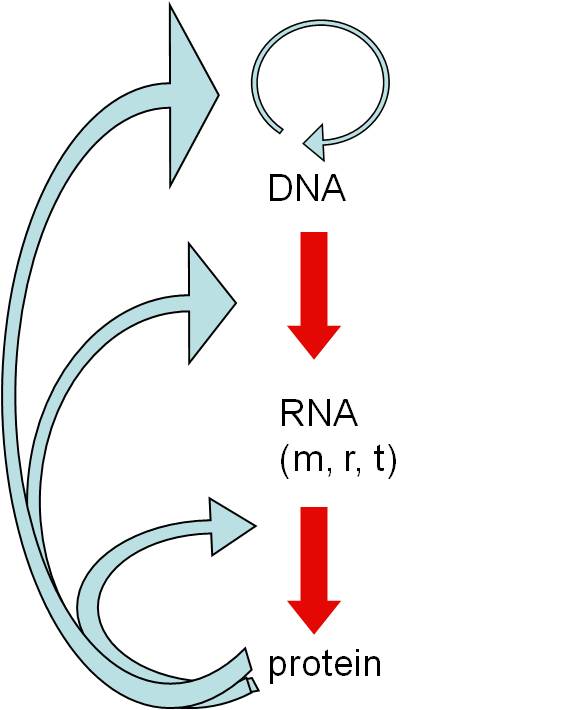 3.
Genetic System:
3.
Genetic System:
a. Problem:
DNA requires protenaceous enzymes to be replicated and read, but proteins require a DNA recipe... neither can function without the other in living cells. Again, we have an apparently "irreducibly complex system" - it all must be there at once to work. You need an information storage molecule (DNA), and a catalyst to access the information (proteins). Both of these functions must be present at the same time for the system to work.b. Possible Solution:
But there is another component of our genetic system....RNA. And we are finding that RNA does lots of wild stuff....
a.Even in current systems, RNA is message (m-RNA), decoder (r-RNA), and transporter (t-RNA).
b. RNA's have been discovered that have catalytic ability (ribozymes), so they can act as an information storage molecule (like DNA) and as a catalyst (like proteins).
c. some RNA molecules can self-replicate.
d. some RNA molecules (mi-RNA, si-RNA) regulate the production of proteins.
So, the possibility exists that genetic systems evolved from self-replicating RNA molecules. Coding for proteins that assist the replication process would be adaptive. And incorporating DNA, which is more stable than RNA, would also be adaptive, producing the system seen today. This idea has been developed into a more formal hypothesis of the evolution of genetic systems, which we will describe next.
c. Hypothesis for the Evolution of a Genetic System:
 - GENETIC SYSTEM #1: Self-Replicating RNA Arises Spontaneously:
- GENETIC SYSTEM #1: Self-Replicating RNA Arises Spontaneously:
- Self-replicating RNA forms by polymerization of spontaneously produced nucleotides.
- It replicates. That's all it has to do to be a successful genetic system...replicate. Consider this: 90% of your DNA does not code for proteins - in some cases it may be regulatory, but there is ALOT of DNA that seems to have no function at all. However, when the DNA is replicated, this non-coding DNA is passed along to the next generation, too. In that sense, it is just as successful as the genes that do code for a protein. So, replication is REALLY all that a genetic system has to do to perpetuate itself through time.
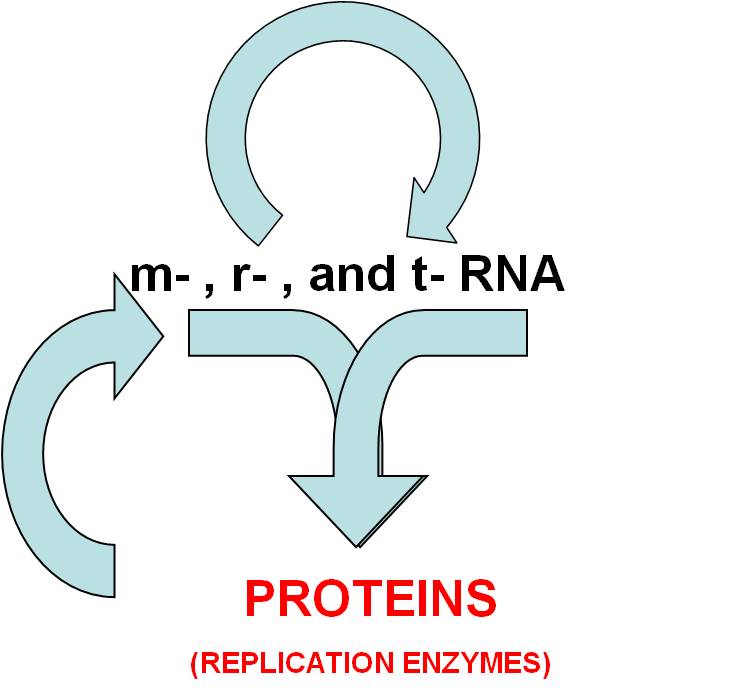 - But, it could replicate more effectively if it coded for replicating proteins.
So, m-RNA and r-RNA and t-RNA interact today to make proteins. If this
step happened next, then the replicating RNA might code for a protein
(an RNA polymerase) that increased the efficiency of its replication.
- But, it could replicate more effectively if it coded for replicating proteins.
So, m-RNA and r-RNA and t-RNA interact today to make proteins. If this
step happened next, then the replicating RNA might code for a protein
(an RNA polymerase) that increased the efficiency of its replication.
- GENETIC SYSTEM #2: RNA systems produces useful replication enzymes.
- So, now there is VARIATION IN GENETIC SYSTEMS... the old, slow, self-replicating RNA, and the new, more rapidly replicating RNA that codes for replication enzymes. So, we have variation and DIFFERENTIAL REPRODUCTIVE SUCCESS... different RNA's that replicate (reproduce) at different rates. What will happen????? Well, the more rapidly replicating molecules will be accumulate over time. They will increase in frequency in the "population" of chemicals...this is selection at a chemical level.
- GENETIC SYSTEM #3: Incorporating DNA
- Now, how and why does DNA get involved? Well, first the how. If the RNA has a small mutation, it may make a slightly different protein... the protein might now link DNA nucleotides together, rather than RNA nucleotides (like the existing RNA replication enzymes). This new enzyme is a 'reverse transcriptase' - it reads RNA and makes DNA. So now, the RNA is read and a DNA molecule is made. (Does this ever really happen today? Are there enzymes that do this? Yes, in retroviruses there are genes that code for reverse transcriptases that copy the viral RNA, make DNA, and then the DNA is inserted into the host's genome. When this viral DNA is transcribed, the new viral genome of RNA is made.)
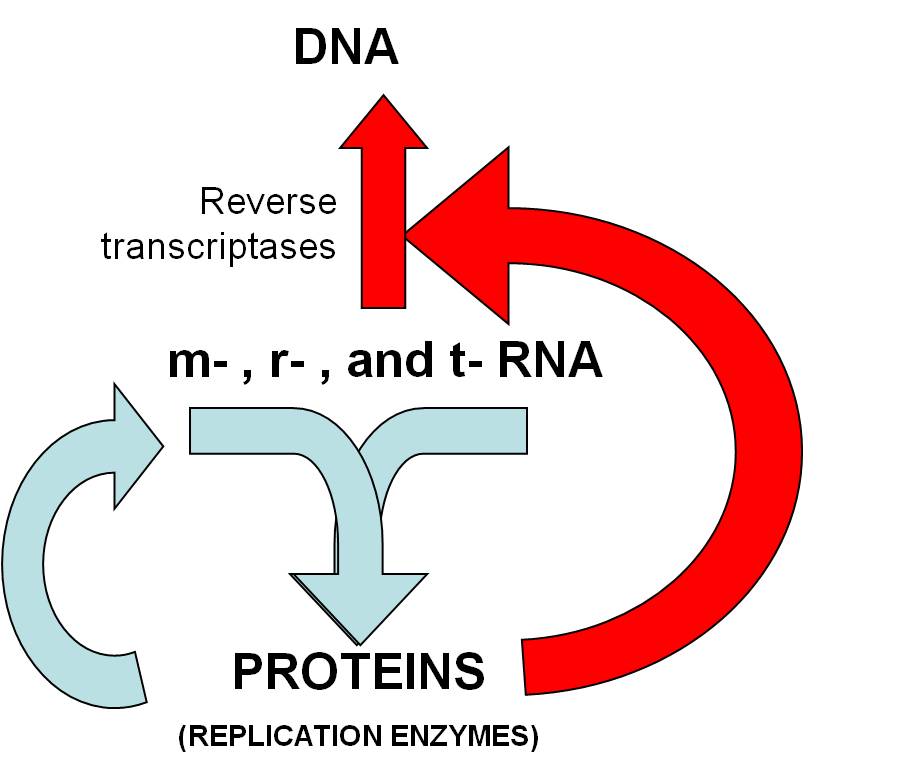 - So, there is now a DNA molecule, and a DNA polymerase that can make a copy
and make it double-stranded. There are also RNA polymerases (present already)
which read a nulcei acid and make RNA. So, here we have evolved a system
where DNA codes for RNA that codes for protein, and proteins are used for DNA
replication, transcription, and translation, JUST LIKE THE GENETIC SYSTEM WE
HAVE TODAY.
- So, there is now a DNA molecule, and a DNA polymerase that can make a copy
and make it double-stranded. There are also RNA polymerases (present already)
which read a nulcei acid and make RNA. So, here we have evolved a system
where DNA codes for RNA that codes for protein, and proteins are used for DNA
replication, transcription, and translation, JUST LIKE THE GENETIC SYSTEM WE
HAVE TODAY.
- Why? Why would DNA be advantageous? Because its' double helix is more stable than the single helices of RNA. Greater stability means fewer mutations, which is a good thing (adaptive). In addition, a two-step process of protein synthesis produces more proteins than a one-step process. Here's the idea in a nutshell. Suppose nucleic acids can be read at a given rate - say, 5 products per second. In a one-step process (RNA coding for proteins), 5 proteins can be made by reading the RNA template each second. So in two seconds, 10 proteins can be made. However, suppose that the first second was used to make copies of the RNA template? In a two-step process of DNA--> RNA--> protein, the first second is used to read the DNA and make 5 products... but these products are 5 RNA molecules. In the next second, each of these 5 RNA's can be used as a template for protein construction, with 5 proteins made off each of them...for a total of 25 proteins after 2 seconds. More productive.
d. How has science tested this hypothesis?
Well, the history goes like this. After the discovery of ribozymes, some scientists saw that RNA could act as both information storage (recipe) AND catalyst (like enzymes). And, since RNA does everything in today's genetic system anyway, the "RNA-first" hypothesis was raised.
So, scientists tested the premise that RNA could self-replicate. They
tested and confirmed this hypothesis - they synthesized molecules that, under
particular 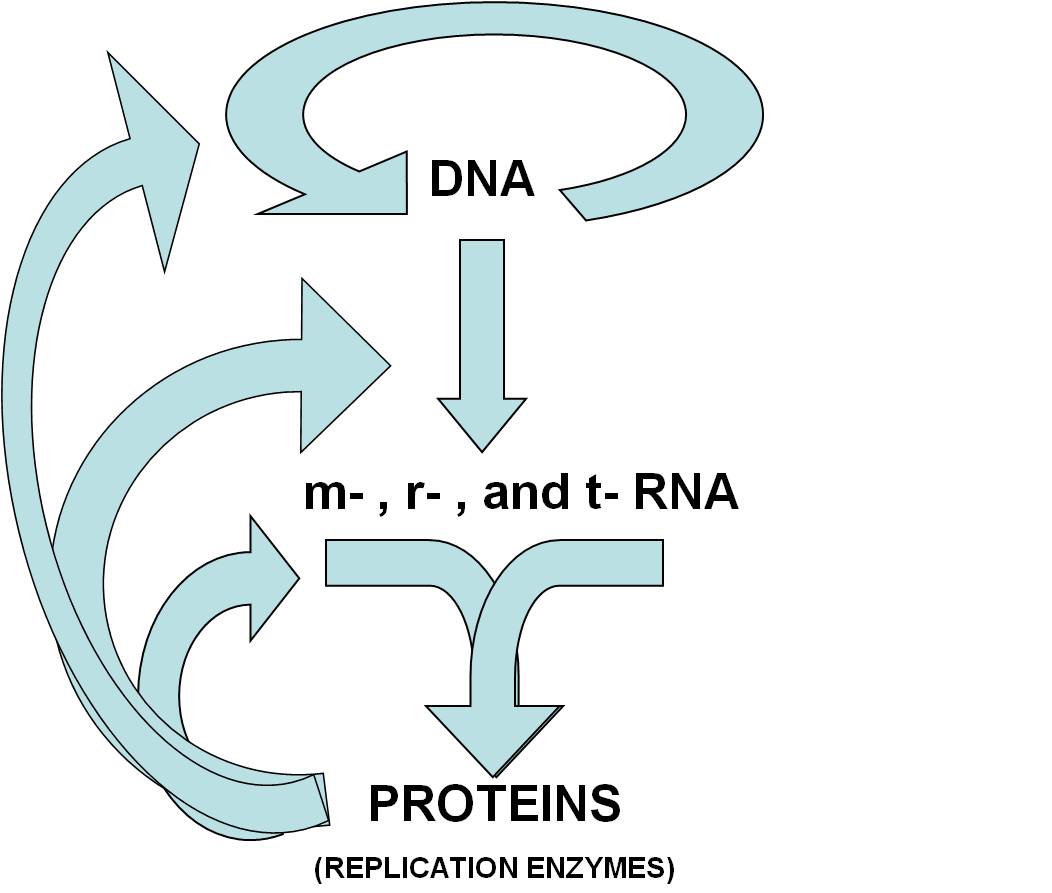 conditions, did self-replicate
(Hypothesis tested and confirmed, in part).
conditions, did self-replicate
(Hypothesis tested and confirmed, in part).
Second, scientists realized that RNA would have to code for DNA to get from SYSTEM #2 to SYSTEM #3. They saw that the existence of retroviruses that confirm this requirement. Retroviruses, like HIV, have an RNA genetic system. They insert this in their host's cell, and this RNA is read like any other m-RNA molecule. One of the genes on the RNA codes for a protein called "reverse transcriptase". This enzyme reads the viral RNA and links together complementary DNA bases - it reads RNA and makes DNA (the reverse of transcription, thus 'reverse transcriptase'). The DNA is then incorporated in the host's genome - it is spliced right into a chromosome. And now, anytime the cell replicates its OWN DNA and divides, the viral gene gets copied and gets passed to the daughter cells. Eventually, some environmental cue stimulates the viral DNA to be read, and its expression (coding for copies and viral replication) kills all the host cells that have descended from this original infected ancestor.
So, parts of this hypothesis have been tested. However, the entire process of evolving a DNA-based genetic system from an RNA-based genetic system has not been done yet. It is certainly intriguing stuff. There is certainly enough evidence, it would seem, to suggest that the hypothesis "IT CAN'T HAPPEN" (such as intelligent designer's claim) needs to be put on hold for a while. Thankfully there are curious people who continue to ask questions.
Scientists continue to find new properties to RNA. Lately (since 2002), a new class of regulatory molecules have been found that can turn genes off. They are short strands of RNA, called "interference-RNA" (mi-RNA and si-RNA). They bind to either DNA or to the m-RNA transcript - they block protein synthesis and effectively turn a gene off. So, RNA not only is a message, a decoder, and a transporter, it is also a genetic regulator. Many medical researchers are trying to use these short pieces of RNA as a blocking agents to stop the progression of genetic diseases.
This pathway is now accepted in the scientific community as a "plausible hypothesis" for the evolution of a genetic system. There is impressive support thus far, but all elements of the hypothesis have not been confirmed. THE SEARCH FOR KNOWLEDGE CONTINUES....!!
D. Summary:
STEPS REQUIRED FOR THE SPONTANEOUS, NATURAL FORMATION OF LIFE, and the evidence to date:
1. Spontaneous synthesis of biomolecules - strong evidence; Miller-Urey experiments.
2. Polymerization of monomers into polymers (proteins, RNA, sugars, fats, etc.) - strong evidence; Fox and Cairns-Smith experiments.
3. Formation of membranes - strong evidence; behavior of phospholipids in solution.
4. Evolution of metabolic systems - reasonable hypotheses, and genetic similarity in genes involved in particular pathways (suggesting gene duplication and susequent evolution of new genes and elaboration of existing pathways)
5. Evolution of a genetic system - a reasonable hypothesis and significant corroborating evidence that it could happen. But no experimental evidence of the process evolving through all three steps.
6. How did these three elements (membrane, metabolism, genetic system come together?) a few untested hypotheses.
So, as you can see, there are several elements of this hypothesis that have various levels of support.
The use of fossils and genetic phylogenies have allowed scientists to reconstruct the history of life and the major epochs and innovations of evolution that have occurred over the last 4 billion years of earth's history. Perhaps the most surprizing thing that we have learned is that we humans are very late arrivals on the evolutionary stage, and that the play had been going on just fine for quite a long while without us. Indeed, the evolution of anything you might recognize as an 'animal' is a fairly recent event in the evolutionary history of the planet; animals have only been present for the last 1/7th of earth history. Although we might think of macroscopic animals and plants as the 'most important' forms of life somehow, it is really the little things - bacteria and archaeans - that keep the biosphere running and allow animals and plants to survive. Without bacteria, you would not digest your food efficiently, plants would have a more difficult time absorbing nutrients, and the nutrients in dead organisms would not be recycled and made available to those of us that are still alive. The basic life processes that drive the biosphere - things like nitrogen fixation, photosynthesis, respiration and energy conversion, and decomposition - are still driven by bacteria. In the first part of this lecture, we'll see that is, indeed, how it has been for the majority of earth history.
 We
have summarized some of the major epochs in the history of the Earth before,
but we will review them here and add in important evolutionary events in the
history of life.
We
have summarized some of the major epochs in the history of the Earth before,
but we will review them here and add in important evolutionary events in the
history of life.
4.5 billion years ago - Earth formed (Evidence: meteorites and moon rocks date to this age, dating the condensation of planetissimals in the solar system)
4.0 bya - oldest rocks (The oldest rocks are not as old as the Earth because the Earth's surface was molten - as a result of constant meteorite bombardment and the energy released during the stratification of Earth's core, mantle, and crust. In addition, the Earth is tectonically active, recycling its crust. So, older rocks that may have solidified may have been subducted and melted)
3.5-3.8 bya - origin of life (The oldest fossils, both microfossils and stromatolites, date to 3.5-3.8 bya). These organisms are similar to prokaryotes we see today. However, it is not possible to distinguish whether they are Bacteria or Archaeans. However, from this period forward there are anaerobic and non-oxygenic photosynthetic bacteria.
2.5-2.0 bya - oxygen accumulating ("red bed" sediments form, demonstrating that oxygen gas in the water was oxidizing iron); this is evidence of the evolution of oxygenic photosynthesis. With the accumulation of oxygen, many obligate anaearobes undoubtedly go extinct. However, other bacteria evolve the capacity to USE oxygen as an oxidizing agent, making the harvest of energy from glucose metabolism more efficient... in other words, aerobic respiration evolves.
2.0 bya - eukaryotes evolve through endosymbiotic absorption/tolerance of aerobic bacteria (protomitochondria). With the evolution of eukaryotes comes the evolution of sexual reproduction. These endosymbionts may have been a cellular parasite, like Rickettsia. Subsequent lines of eukaryotes absorbed photosynthetic cyanobacteria, which evolved into symbiotic chloroplasts. The evidence for these theories of eukaryote evolution are the genetic similarity between mitochondria and chloroplasts (that have their own DNA) and various types of bacteria and cyanobacteria. So until this point - representing nearly 1/2 of life's history on this planet - life was exclusively bacterial. The major types of ecological roles, from nitrogen fixation, to photosynthesis, to heterotrophy, to saprophagy (eating dead stuff) all evolved among bacteria - creating food webs and biogeochemical cycles that shunted energy through the biosphere and other earth systems.
0.7 bya - the first animals evolve - sponges and then soft-bodies invertebrate fossils, often radially symmetrical like jellyfish.
0.5 bya - radiation of animals with hard bodies (Cambrian explosion) - beginning of the Paleozoic Era ('old animals')
0.4 bya - evolution of land animals and plants
0.24 bya (240 mya) - Permian extinction; end of the Paleozoic Era
240-65 mya - Mesozoic Era (Age of Dinosaurs - 'middle animals')
65 mya - Cretaceous extinction (meteorite impact off Yucatan Peninsula) - beginning of the 'Cenozoic Era' ('recent animals')
6-5 mya - split of chimp and human lineages
4.5 mya - Australopithecines (1/1000 of earth's history left to present...99.9 % has occurred already).
0.2 mya (200,000) - oldest Homo sapiens fossil. (99.995 % of earth's history has passed).
The Linnaean system is a heirarchical system that places species within genera, and similar genera within higher nested categories. Historically, these determinations were based on similarities in morphology. There are lots of mnemonics for remembering the Linnaean classification system: "Dandy King Philip Came Over From Great Spain" is one way to remember the sequence of "Domain, Kingdom, Phylum, Class, Order, Family, Genus, and Species". However, it doesn't work for plants, in which 'Division' is used instead of phylum. The Linnaean system lended itself to evolutionary reinterpretations, because evolution also produces nested patterns of relationships. The process of divergence creates an evolutionary 'tree', and each twig (species) is nested with other small twigs as part of one branch, and several branches form one limb, and the limbs unite to form the trunk.
Although Linnaean classification and evolution both produce nested heirarchies, they are not necessarily equivalent. For instance, if we have multiple genera within a family, the Linnaean system does not necessarily describe the relationships among these genera; all it describes is their mutual presence in this one family. For example, in the figure below left, the Linnaean systems allocates cats into four genera, but does not describe the relationships among these genera. However, evolution - through phylogenetic systematics - tries to determine which taxa (species, genera, families, etc.) are more closely related and share a more recent common ancestor. Indeed, phylogenetic systematics would argue that the degree of biological relatedness among taxa should be reflected in their taxonomic affinity. This becomes a goal of phylogenetic systematics - to create a taxonomy that represents the evolutionary relationships among species, genera, families, etc. The goal is to create a monophyletic taxonomy that include all organisms decended from a common ancestor.
 |
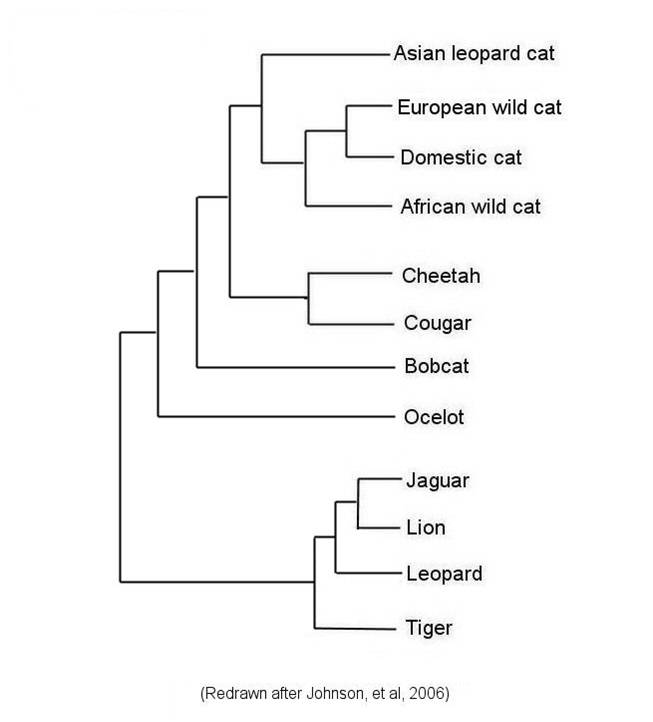 |
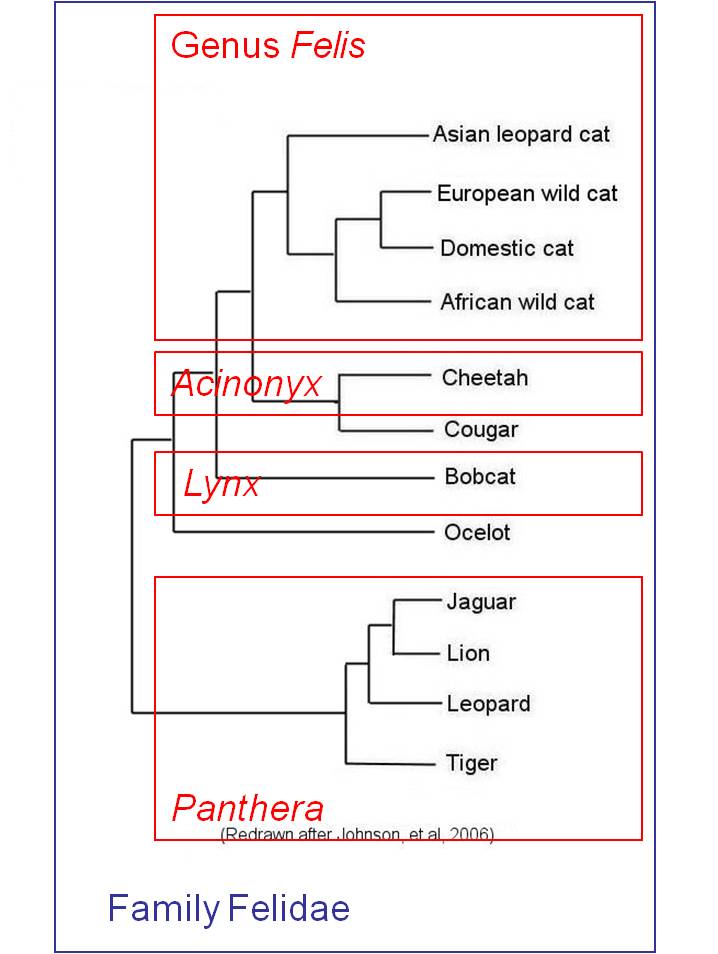 |
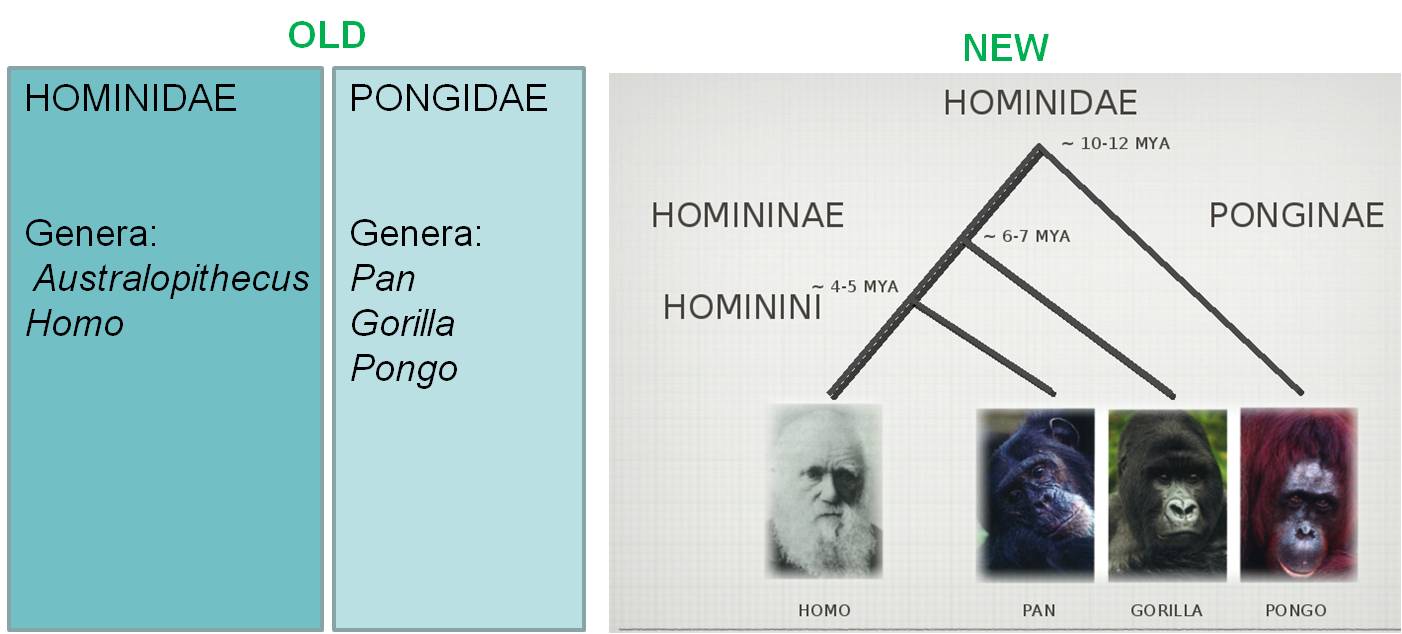 The
Linnaean system was based on morphology, alone, and we know that morphology
can sometimes misrepresent patterns of genetic relatedness (remember sibling
species and polymorphic species?). So, as we learn more about the degree of
genetic relatedness among species, we occasionally find that some species that
are closely related have been placed in different taxa. For example, the historical
classification of humans, chimps, gorillas, and orangutans placed humans in
the family Hominidae, and the other three species in the family Pongidae. This
does not represent their degree of biological relatedness, however, because
human and chimps share a more recent common ancestor with one another. So, to
include all descendants of a common ancestor (and that ancestor), chimps and
humans should be placed in the same taxonomic group (either genus or family).
This could include gorillas (as all three share a common ancestor), or could
also include gorillas and orangutans (as all four share a common ancestor),
but can't split humans and chimps into separate groups.
The
Linnaean system was based on morphology, alone, and we know that morphology
can sometimes misrepresent patterns of genetic relatedness (remember sibling
species and polymorphic species?). So, as we learn more about the degree of
genetic relatedness among species, we occasionally find that some species that
are closely related have been placed in different taxa. For example, the historical
classification of humans, chimps, gorillas, and orangutans placed humans in
the family Hominidae, and the other three species in the family Pongidae. This
does not represent their degree of biological relatedness, however, because
human and chimps share a more recent common ancestor with one another. So, to
include all descendants of a common ancestor (and that ancestor), chimps and
humans should be placed in the same taxonomic group (either genus or family).
This could include gorillas (as all three share a common ancestor), or could
also include gorillas and orangutans (as all four share a common ancestor),
but can't split humans and chimps into separate groups.
Another classic example is the treatment of mammals, birds, and reptiles. In Linnaean taxonomy, these three groups of animals are placed in different Classes: Mammalia, Reptilia, and Aves (birds). But phylogenetic systematics groups them differently (see the tree of life site). Some reptilian groups, like extinct dinosaurs and extant crocodilians, are more closely related to birds than they are to other reptiles like lizards, snakes, and turtles. And some "reptiles", like the sail-finned Pelycosaurs, are more closely related to mammals than to other reptiles. So, phylogenetic systematics separates the "synapsids" (mammals and mammal-like reptiles), from "reptilia", and divides reptilia into the "anapsids" (turtles) and "diapsids". The diapsids are further divided into the "archosauromorphs" (crocodilians, birds, and extinct dinosaur relatives) and the "lepidosauromorphs" (modern lizards ands snakes, and their extinct ancestors). Because the number of clades does not conform to the number of taxonomic levels, the notion of "class" and "order" becomes problematic. However, knowing that "archosauromorphs" contains croc and birds but not turtles, lizards, or snakes correctly represents the evolutionary relationships among these groups, and conveys more correct evolutionary information than the term 'reptile'.
Finally, a third example highlights the issue at the largest scale of taxonomic resolution, and brings us back to bacteria. Prior to the 1980's, most biologists classified all prokaryotes in the Kingdom Monera. While they recognized that there was great diversity in the group, biologists considered the absence of a nucleus a fundamental, distinguishing morphological feature of the group that separated it from all eukaryotes. However, genetic analyses in the 1980's by Carl Woese suggested that there were two major clades of prokaryotes - the Bacteria and the Archaea. And actually, most genetic analyses show that the Archaea are more closely related to the eukaryotes than to the bacteria. So, using the criteria of phylogenetic systematics, it is improper to classify all prokaryotes together. Rather, Woese suggested that a new level be created - the domain, and that life be split into three domains: the Bacteria, the Archaea, and the Eukarya.
III. The Prokaryote Domains: Bacteria and Archaea
A. Overview
 These
two domains are ancient; they may even have diverged before 3.5 bya. The problem
with using genetic clocks and genetic similarity to attempt to date this point
of genetic divergence on these groups is that prokaryotes exchange DNA "laterally".
They don't just pass it down a lineage from parental cell to daughter cells;
they can also pass genes from one living bacterium to another through conjugation
or transformation. As such, genetic similarity can occur as a result of lateral
gene transfer between unrelated organisms, and thus the degree of genetic similarity
in particular genes does not necessarily indicate the degree of common ancestry.
For these reasons, it is very difficult to use this particular technique to
date the time of divergence of these two ancient groups.
These
two domains are ancient; they may even have diverged before 3.5 bya. The problem
with using genetic clocks and genetic similarity to attempt to date this point
of genetic divergence on these groups is that prokaryotes exchange DNA "laterally".
They don't just pass it down a lineage from parental cell to daughter cells;
they can also pass genes from one living bacterium to another through conjugation
or transformation. As such, genetic similarity can occur as a result of lateral
gene transfer between unrelated organisms, and thus the degree of genetic similarity
in particular genes does not necessarily indicate the degree of common ancestry.
For these reasons, it is very difficult to use this particular technique to
date the time of divergence of these two ancient groups.
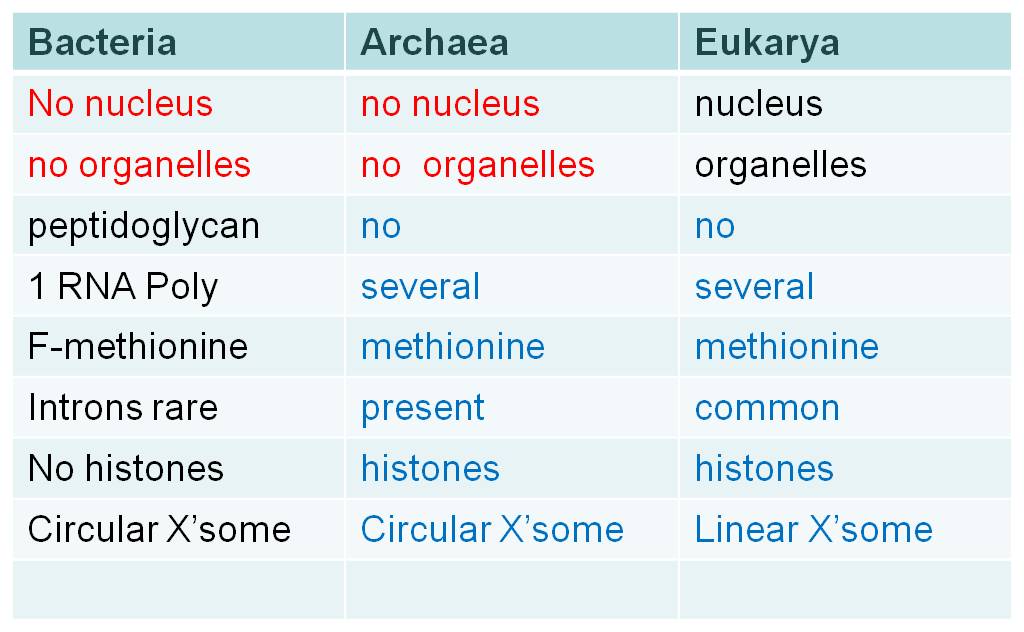
Although they share some fundamental similarities like the lack of a nucleus, the lack of membrane-bound organelles, and circular chromosomes, these two prokaryotic domains differ in some important respects. When they differ, the archaeans are often similar to the Eukarya. For example, bacteria have peptidoglycan in their cell walls, while archaeans (and eukaryotes) do not. Bacteria have only one type of RNA Polymerase; archaeans (and eukaryotes) have several. Bacteria use formyl-methionine as the initiator amino acid in translation, archaeans (and eukaryotes) use methionine. Histones are not associated with bacterial chromosomes, but they are associated with archaean (and eukaryote) chromosomes. Introns are rare in bacterial genes, but are progressively more common in archaeans and eukaryotes. So, there are several fundamental differences between these two groups, and often the archaeans are more similar to eukaryotes than to bacteria.
1. Archaea
 The
archaea are very diverse, and include groups that have adapted to some of the
most inhospitable environments on Earth. Indeed, many of these environments
may be similar to those that were common in early earth history. So, rather
than thinking that these organisms have evolved to exploit these isolated odd
habitats, perhaps they adapted to these conditions when the conditions were
common, and have just 'hung on' in these habitats that have become rare over
time as the earth has changed. The extreme thermophiles live at very high temperatures at sulphur springs and geothermal vents where
temperatures can exceed 1000C. How they protect their proteins and
DNA under these conditions is an area of active research. Another group are
tolerant of very extreme salt concentrations - these are the extreme halophiles.
Yet another group of extreme organisms are the methanogens. These organisms
use H2 to reduce CO2 and produce energy; producing CH4 (methane or 'swamp gas') as a waste product. Most are obligate anaerobes, meaning
that they can only survive in environments where the concentration of oxygen
gas is very low. However, some are aerotolerant, and can survive in the presence
of oxygen though they do not use it for respiration. These conditions occur
at the bottom of deep water bodies, or where the action of respiring bacteria
deplete available oxygen - like in swamp and marsh sediments and in the digestive
tract of animals (including humans). Some methanogens are extreme thermophiles,
too.
The
archaea are very diverse, and include groups that have adapted to some of the
most inhospitable environments on Earth. Indeed, many of these environments
may be similar to those that were common in early earth history. So, rather
than thinking that these organisms have evolved to exploit these isolated odd
habitats, perhaps they adapted to these conditions when the conditions were
common, and have just 'hung on' in these habitats that have become rare over
time as the earth has changed. The extreme thermophiles live at very high temperatures at sulphur springs and geothermal vents where
temperatures can exceed 1000C. How they protect their proteins and
DNA under these conditions is an area of active research. Another group are
tolerant of very extreme salt concentrations - these are the extreme halophiles.
Yet another group of extreme organisms are the methanogens. These organisms
use H2 to reduce CO2 and produce energy; producing CH4 (methane or 'swamp gas') as a waste product. Most are obligate anaerobes, meaning
that they can only survive in environments where the concentration of oxygen
gas is very low. However, some are aerotolerant, and can survive in the presence
of oxygen though they do not use it for respiration. These conditions occur
at the bottom of deep water bodies, or where the action of respiring bacteria
deplete available oxygen - like in swamp and marsh sediments and in the digestive
tract of animals (including humans). Some methanogens are extreme thermophiles,
too.
Although the Archaea are noted for representatives that tolerate these extreme environments, there are other archaeans that live under more benign environmental conditions across our planet, where they function as photosynthetic producers, consumers, and decomposers. Curiously, although archaeans do inhabit the human gut, no archaeans have been identified that cause human disease. The other prokaryotes - the bacteria - contain many species that cause human disease.
2. Bacteria
 The
bacteria are as diverse than the archaeans, expressing an extraordinary variety
of metabolic processes. A couple notable groups are the cyanobacteria, thought
to share common ancestry with eukaryotic chloroplasts, and the gram-positive
bacteria that have many types of pathogenic bacteria, such as those causing
staph and strep infections, botulism, tuberculosis, anthrax, and many other
diseases.
The
bacteria are as diverse than the archaeans, expressing an extraordinary variety
of metabolic processes. A couple notable groups are the cyanobacteria, thought
to share common ancestry with eukaryotic chloroplasts, and the gram-positive
bacteria that have many types of pathogenic bacteria, such as those causing
staph and strep infections, botulism, tuberculosis, anthrax, and many other
diseases.
Both the bacteria and archaea are very old groups, and have been radiating and evolving for over 3.0 billion years. Recent genetic studies have revealed an astounding degree of genetic variation within these groups - an understandable consequence of their long evolutionary histories. This genetic variation is reflected by the 'branch lengths' in the tree diagram to the right. The genetic variation among the plants, animals, and fungi, represented by their relative short branch lengths, are dwarfed by the genetic variation among the archean and bacterial clades. But of course, the animals have only been radiating for the last 0.7 billion years. So, the archean and bacterial lineages have been radiating for 5 times as long and so it is not surprizing that they are much more diverse. Thus, as expected for groups that are so old, they have radiated dramatically - even within subgroups. For this reason, even groups that share common ancestry can include organisms that are very different. For our purposes here, an overview of the diversity is presented, rather than a overview of their classification or phylogeny.
If we examine the morphological diversity in prokaryotes, we find round (coccus), rod-shaped (bacillus), and spiral (spirochaete). That is not a lot of morphological diversity! So how have the prokaryotes been changing over the last 3.5 by? Well, they have been evolving metabolically. The theme of bacteria diversity is metabolic diversity. There are very few metabolic processes that eukaryotes can perform that prokaryotes cannot. However, the reverse is not true; the prokayotes have a wide range of metabolic diversity - a range that, in many respects, exceeds the metabolic diversity of eukaryotes.
1. Requirements of Oxygen for Cellular Respiration
Consider types of respiration. There
are no anaerobic eukaryotes. Most eukaryotes require oxygen (they have mitochondria,
after all), although some can function without it for a short time (facultative
anaerobes like yeast). Prokaryotes, on the other hand, have a wide range of
'respiratory' strategies:
- some bacteria, like methanogens, cannot live in the presence of oxygen - obligate
anaerobes
- some bacteria can tolerate oxygen
but don't use it - aerotolerant
- some bacteria can live with or without oxygen - facultative aerobes
- some bacteria live only in the presence of oxygen - obilgate aerobes
2. Ecological Roles
- primary producers: bacteria still make a significant contribution to the absorption of CO2 and the production of oxygen on our planet - largely in the oceans. And these oceanic bacteria are the bottom of marine food chains.
- nitrogen-fixing bacteria: can convert N2 into NH3 (ammonia) and then NO3 (nitrates) which can be taken up by plants. Only bacteria can do this; and so all new, natural inputs of nitrogen to the biosphere are by bacteria. (Lots of biologically accessible nitrogen is added to the environment by humans - through fertilizer and the production of NOx compounds by internal combustion engines).
- decomposers: as consumers of dead organic matter, bacteria (and fungi) break down complex organics and harvest energy. And the byproducts of that digestion are mineral nutrients (N, P, K) that are made available for reuptake in the biosphere by plants. So, bacteria are needed for nutrients to cycle through ecosystems.
- pathogenic bacteria: these bacteria get energy from living tissue and destroy cells in the process; they also produce either exotoxins that they secrete, or endotoxins that are bound to the cell wall. Examples of these pathogenic bacterial diseases are: staph infections, salmonella, typhoid, cholera, scarlet fever, tetanus, tuberculosis, botulism, gonorrhea, and syphilis.
- symbiotes: there are gut bacteria in nearly all animals that assist with digestion and may be required for the production and absorption of certain metabolites.There are also bacterial symbionts in plants, protists, and fungi.
As such, bacteria interact in every way possible with other organisms, and form the 'hub' of the biosphere. In some ways, the evolution of life has been an "embellishment" of the bacterial ecosystems that dominated the planet for the first 2.3 billion years of life.
The take home message about prokaryotes is their metabolic diversity. For 1.8 billion years, life on Earth was SOLELY bacterial. How was it evolving? How was it responding to the environment? Well, it seems that life was responding metabolically, adapting by exploiting different environments and energy sources in different ways.
The eukaryotes have been evolving for at least 2.0 billion years. As you might expect (as was argued for the prokaryotes groups), there has been considerable divergence within this group, producing a dramatic range of genetic and metabolic variation. And of course, the multicellular eukaryotes evolved from protistan ancestors - so multicellular eukaryotes (plants, fungi, and animals) are included within this clade. So, the focus will be on the origin of the clade (Eukarya) and the diversity within this clade. We will pay particular attention to the sister groups of the multicellular "kingdoms" that we will consider in greater detail in the remaining lectures. Obviously, the features that distinguish eukaryotes from archaean ancestors are the presence of membrane-bound organelles (mitochondria and chloroplasts) and a nucleus. So, any hypothesis of origin must explain how these defining structures came into existence.
1. Origin of Organelles
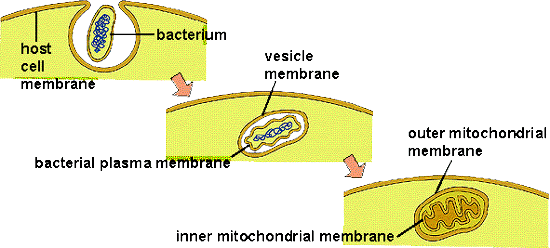
The origin of eukaryotic cells is described by the Endosymbiotic Theory, popularized by Lynn Margulis in the 1960's. It states that organelles are descended from free-living bacterial cells that were absorbed by - or parasitized - archaeans. The evidence for this idea is very strong. Both mitochondria and chloroplasts have double membranes. The outer membranes are similar to the plasma membrane of eukaryotes and archaeans. The inner membrane has certain attributes of bacteria. If a bacterium were absorbed through the process of endocytosis, the bacterium would be surrounded by the membrane of the host's vacuole - creating a double membrane. In addition, both types of organelles have their own DNA, which is circular like bacterial DNA. Both mitochondria and chloroplasts divide by fission, independent of the mitotic division of the eukaryotic cell they inhabit. Over the course of evolutionary history, genes have been exchanged between the organelles and the nucleus, or have been eliminated in the organelle because they also exist in the nucleus. For these reasons, the organelles are functionally dependent on the eukaryotic cell and can no longer live independently for very long.
Additional similarities link these organelles to specific bacteria. Mitochondria not only have their own DNA, but they also produce their own proteins using these DNA recipes. (That should be no surprize!) Protein synthesis occurs on ribosomes within the mitochondria, and their ribosomes are more similar to bacterial ribosomes than to the ribosomes in the cytoplasm of their eukaryotic host cells (produced by the nuclear DNA). Likewise, the genetic code used by mitochondria has a few differences from the universal genetic code; differences also seen in the proteobacteria. Genetic analyses suggest that mitochondria are most closely related to Rickettsia proteobacteria. Rickettsia does not causes "rickets" (a vitamin-D deficiency); rather, Rickettsia bacteria cause a variety of human diseases like many forms of tick-borne fevers (Rocky Mountain spotted fever) and typhus. In addition to having a similar genome to mitochondria, these bacteria are obligate intracellular parasites - so they move into eukaryotic cells and become "organelle-like". In addition, Rickettsia produce ATP in the same way that mitochondria do. So, it is possible that archaeans absorbed bacteria and used them for ATP production, or that the bacteria infected the archaeans to consume their sugars, or some combination of these scenarios. In any case, the bacteria and archaean ultimately benefitted from the interaction, forming a mutualistic or "symbiotic" relationship.
 |
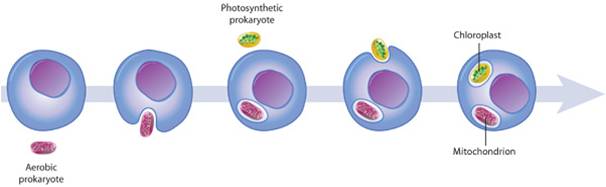 |
Chloroplasts have an independent, endosymbiotic origin; they are genetically similar to a different group of bacteria called cyanobacteria. Some have a double membrane like mitochondria. However, in some protistan groups, the chloroplasts have 3 or 4 membranes, suggesting multiple rounds of endosymbiosis. So, the current model of chloroplast ancestry suggests a single origin of photosynthetic red and green algae. Green algae were then absorbed (secondary endosymbiosis) by other unrelated protists, creating the euglenids and chlorarachniophytes. These organisms are not related directly through nuclear genetic analysis, but they have very similar, multi-membraned chloroplasts - suggesting independent endocytosis of green algae. A similar process may explain the chloroplast similarity in the stramenopiles, apicomplexans and dinoflagellates - independent endocytosis of a red algae could explain their dissimilar nuclear DNA and their similar, multi-membraned chloroplasts.

There are several groups of eukarya; most of them contain only single-celled representatives.
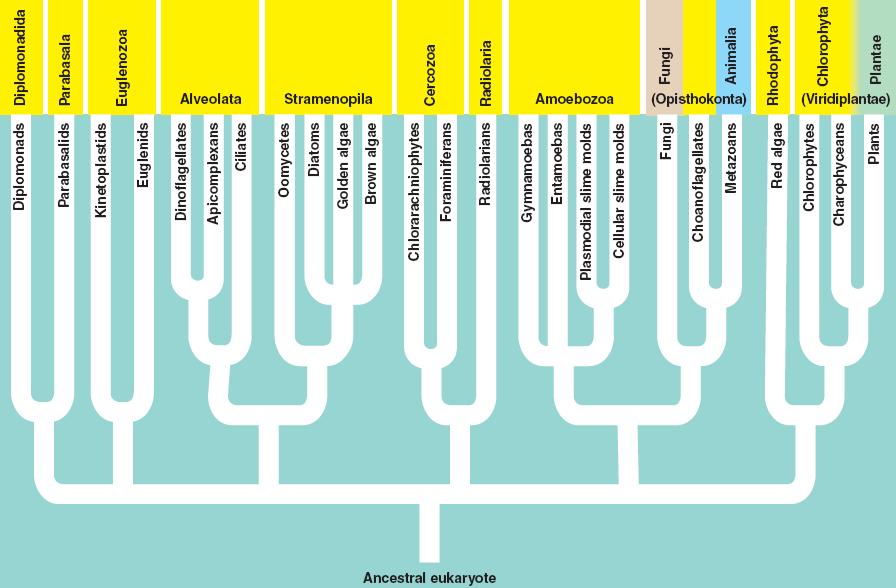
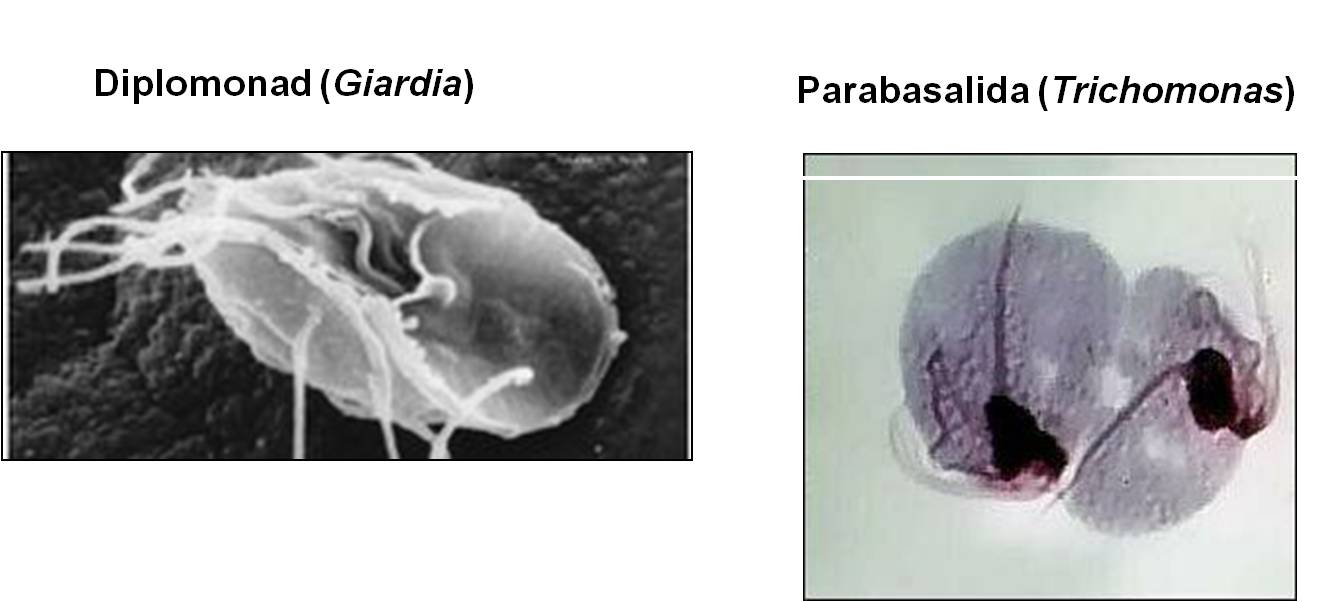 1.
Diplomonada and Parabasalida
1.
Diplomonada and Parabasalida
These two groups are unusual in having very reduced (even absent) mitochondria. 'Reduced' means that they do not perform all of the functions of mitochondria found in other eukaryotes. Although some scientisits initially thought that these might be transitional forms between archaeans and eukaryotes, the thought now is that these groups have lost mitochondrial function because they have adapted to anaerobic environments. Without oxygen present, the mitochondria becomes a non-functional, vestigial organelle. Giardia is a diplomonad (two equal nuclei) and Trichomonas is a Parabasalid. Giardia is an intestinal parasite of vertebrates, and can infect the human gut where it causes acute diarrhea, gas, bloating, and dyehydration and weight loss. Trichomonas vaginalis infects the vagina and urogenital tract; it is a common sexually transmitted disease that usually causes no symptoms in males but does cause itching, discomfort, and discharge in females. This infection is called trichomoniasis or 'trich'.
2. Euglenozoa
This group includes the euglenids and the kinetoplastids - two groups in which the cells have a supportive rod in the flagella. Sleeping sickness is caused by members of this group, the Trypanosomes. The euglenids are unusual in being heterophototrophs. They can synthesize their own sugars through photosynthesis, but they can also prey on small prey items. Their chloroplasts have three membranes and are believed to have resulted from secondary endosymbiosis of green algae.
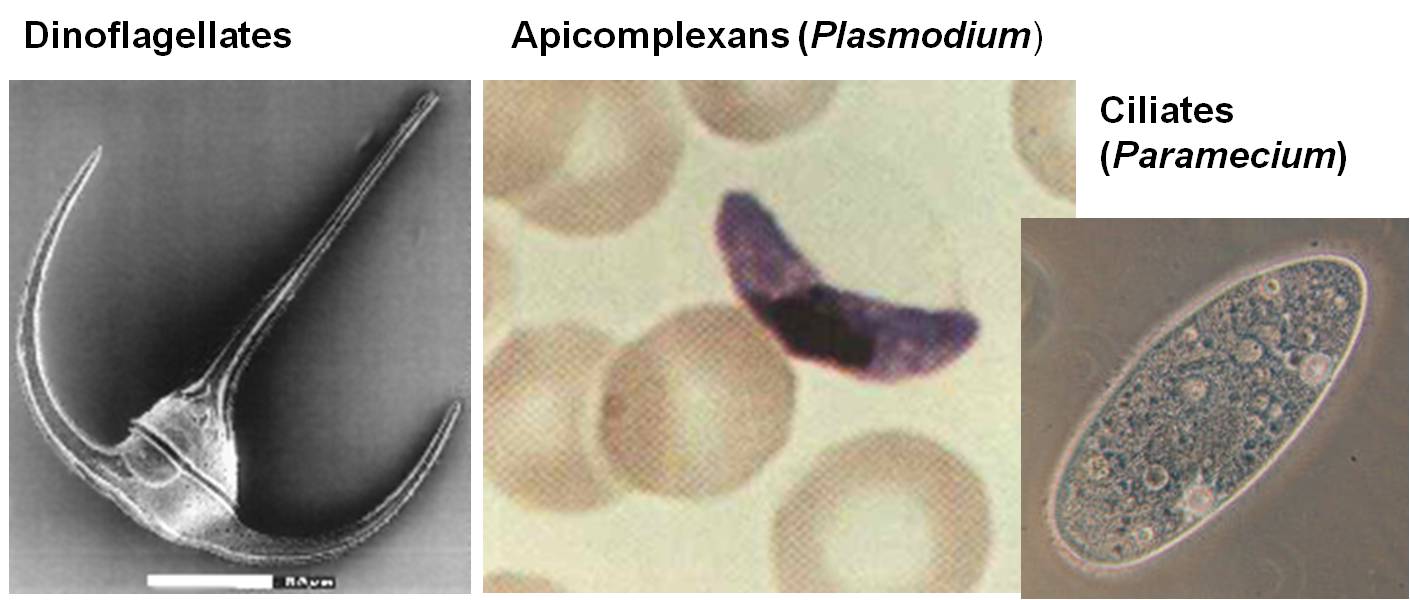 3.
Alveolata
3.
Alveolata
This group of diverse organisms have membrane-bound vesicles nested beneath the plasma membrane. It includes a wide variety of photosynthetic, parasitic, and predatory organisms, including ciliates like Paramecium, Dinoflagellates (cause of 'red tides'), and Apicomplexans like the malarial parasites in the genus Plasmodium.
4. Stramenopiles
This is another diverse group of photosynthetic and heterotrophic organisms, characterized by their two flagella being 'hairy'. Diatoms, golden algae, and brown algae are photosynthetic members of this group. Kelp is a huge multicellular brown algae. They have chloroplasts with four membranes that are thought to have arisen by secondary symbiosis of a red algae.
5. Rhizaria
These sister taxa have thin, threadlike pseudopods that they extend through a shell or 'test'. The Foraminiferans, belong to the Cercozoa group, often harbor symbiotic algae and feed off the sugars these algae produce. By contrast, most radiolarians are predatory, feeding on bacteria and protists that get 'stuck' to these sticky cellular extensions. Deep sedimentary deposits of fossil foraminiferan and radiolarian tests have been found, and are often used as bioindicators that specify strata of a particular age.
6. Amoebozoa
This clade includes organisms with broad pseudopods that are used for locomotion and prey capture. Most amoebozoans are free-living, living in moist soil or freshwater environments. However, the Entamoebas are parasitic, infecting the digestive tract (where they cause amoebic dysentery) or the brain (amoebic meningitis) - both of which can be fatal. Slime molds are also included in this clade. Although they were once thought to be fungal, they do have an amoeboid stage that confirms their genetic relationship to this group. Some slime molds have a strange life cycle in which free-living, independent amoebas aggregate and form a spore-producing structure. Some cells in the mass dry out and become a rigid stalk, while other cells migrate to the end of the stalk and develop into encapsulated spores. New amoebae 'hatch" from the dispersed spores when conditions are favorable. This is an interesting case where protist cells cooperate to form a multicelluar aggregate with the rudiments of cell specialization. However, this is not the group from which multicellular animals or fungi evolved.
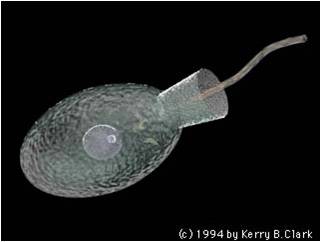 7.
Opisthokonta
7.
Opisthokonta
This clade includes the fungi and animals, and the flagellated sister taxa that bear the stamp of their close phylogenetic relatedness. Other groups have flagella, like the diplomonads and the euglenozoa - but those groups have two flagella and they are at the 'front' of the organism, pulling the organism along surfaces or through a liquid medium. The Opisthokonts, however, have a single, posterior flagellum at some point in the life cycle of the organism. In the animals, this is represented by sperm cells. most fungi do not have flagellated cells, except for the most promitive group, the chytrids. This morphological characteristic is the only morphological trait that links these groups, but their genetic similarity testifies to their common ancestry.
An important opisthokont group besides the fungi and animals are the Choanoflagellates. This group is considered to be the "sister group" to the animals, and may represent what the ancestral animals evolved from. The Choanoflagellates are distinguished by a ring of small projections that ring the flagellum like a collar. These microvilli act like a sieve. The motion of the flagellum wafts bacteria into the ring of microvilli, where bacteria are trapped and then absorbed by phagocytosis. There are over 100 species of choanoflagellates; some are solitary, but some form colonies. Their predation of bacteria provides a valuable link in marine food webs. As we will see when we get to animals, the most primitive animals - the sponges - have cells that are very similar to these choanoflagellates.
 8.
Archaeplastida
8.
Archaeplastida
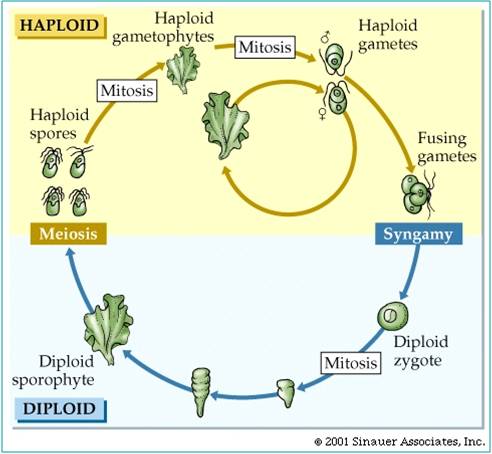
The last eukaryotes clade contains red algae, green algae, and land plants. Red algae have chloroplasts that contain chlorophyll a like cyanobacteria; green algae and plants contain chloroplasts with chlorophyll a and chlorophyll b. The red algae are largely multicellular, colonial organisms best represented by red seaweeds. Green algae are a diverse group, including unicellular, colonial, and multicellular species like Ulva and Enteromorpha (green seaweeds), Volvox, and Spirogyra. A key shared derived character that is seen in the Archaeplastida is "alternation of generation". Consider a haploid organism about to reproduce. This organism produces gametes (egg and/or sperm) by mitosis. These gametes fuse to form a diploid zygote, which developes into the diploid offspring. The diploid organism produces haploid spores by meiosis. These spores do not fertilize one another. Rather, they each divide by mitosis and develop into separate haploid organisms. So, the ploidy level alternates each generation. All plants share this life cycle, so we will see it often during the plant diversity lectures.
Study Questions:
1. How old is the Earth, how old are the oldest rocks on the Earth, and how old are the oldest fossils on Earth?
2. What were the major gases in the Earth’s early atmosphere? Where did it come from?
3. When did oxygen begin to accumulate in the Earth’s oceans? Where did it come from? What geological evidence do we have of this change in ocean chemistry?
4. How could a membrane evolve?
5. What two attributes of RNA lead biochemists to hypothesize that RNA may have been the first genetic system?
6. How and why might the synthesis of proteins by RNA have evolved?
7. How and why might have DNA been incorporated into this process?
8. How do Linnaean classification and cladistic phylogenetic analyses differ?
9. How has the way we classify human changed as a consequence of genetic analyses?
10. What are the three domains of life?
11. Why are aarchaeans and bacteria so much more genetically diverse than plants, animals, and fungi?
12. All eukaryotes are aerobic. List four types of metabolism that prokaryotes have with respect to oxygen.
13. Describe five ecological roles that prokaryotes play in the environment.
14. What characteristics distinguish the Domain Eukarya?
15. Describe the endosymbiotic theory of organelle origin, and provide evidence for this theory.