Evolution 1: How Does Life Change?
I. The Growth of Biological Thought
1. Observations
a. The Earth is old: James
Hutton (1726-1797): Hutton was the first great british geologist.
He compared Hadrian's wall - which looks new but was 1600 years old (122 AD)
- with natural rock outcrops that were strongly weathered. Hutton concluded
that the natural outcrops must be 100's of times older. He also examined an
important formation at Siccar Point, where one series of nearly vertical strata
is overlain by another series of horizontal strata. This is now called an 'unconformity',
and Hutton explained it as follows. Based on Steno's laws of superposition,
the bottom vertical sediments must have been laid down first, and they must
have been laid down horizontally. Ages must have passed between each deposit,
as each turned to rock. Then, uplifts must have occurred to bend them into a
vertical aspect. Long periods of erosion must take place to wear that uplift
flat, followed by the long intervals of time needed to deposit the second horizontal
series. Also, if erosion and deposition acted slowly (as current observations
show), then it must have taken a really long time to erode mountains or build
up marine deposits (White Cliffs of Dover). He concluded that this slow, 'uniformitarian'
cycle of deposition, uplift, erosion, and deposition meant that the Earth was
unfathomably old. Indeed, the cycle may mean that it's age might not be discoverable.
In short, Hutton concludes, the Earth has "no vestige of a beginning, no prospect
of an end."
b. Paleontology: Fossils show Patterns
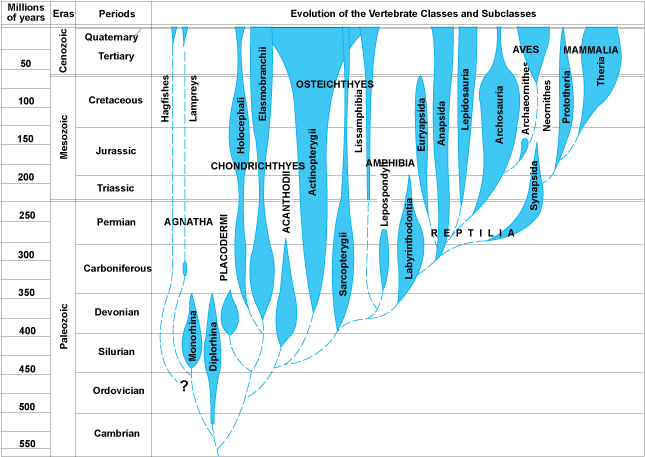 Paleontology
provided a variety of interesting patterns. First, there were extinct forms
that were different from the species alive today. Although some earlier natural
philosophers suggested that the creatures might still exist in some unexplored
corner of the globe, that was a less satisfying hypothesis in the mid-1800's...
most areas of the globe had been visited by Europeans. Also, the idea of extinction
was repugnant to some people on theological grounds. If God had created a perfect
world, then extinction renders that creation imperfect. Also, if species could
go extinct since the creation, could species also come into existence since
the creation? Just how dynamic was this system?
Paleontology
provided a variety of interesting patterns. First, there were extinct forms
that were different from the species alive today. Although some earlier natural
philosophers suggested that the creatures might still exist in some unexplored
corner of the globe, that was a less satisfying hypothesis in the mid-1800's...
most areas of the globe had been visited by Europeans. Also, the idea of extinction
was repugnant to some people on theological grounds. If God had created a perfect
world, then extinction renders that creation imperfect. Also, if species could
go extinct since the creation, could species also come into existence since
the creation? Just how dynamic was this system?
Darwin was impressed by two major
patterns in the fossil record.
1. The major groups of animals accumulate
in an orderly manner'. Everything is not represented at the beginning. In vertebrates,
for instance, the fishes appear first, and exist throughout the rest of the
record. Amphibians appear next, followed by reptiles, mammals, and birds. So
it is not everything at the beginning, and it is not a replacement. Where did
mammals come from? Spontaneous generation had been refuted, so Darwin knew that
mammals had to come from other pre-existing animals. But the only completely
terrestrial vertebrates before mammals were reptiles.
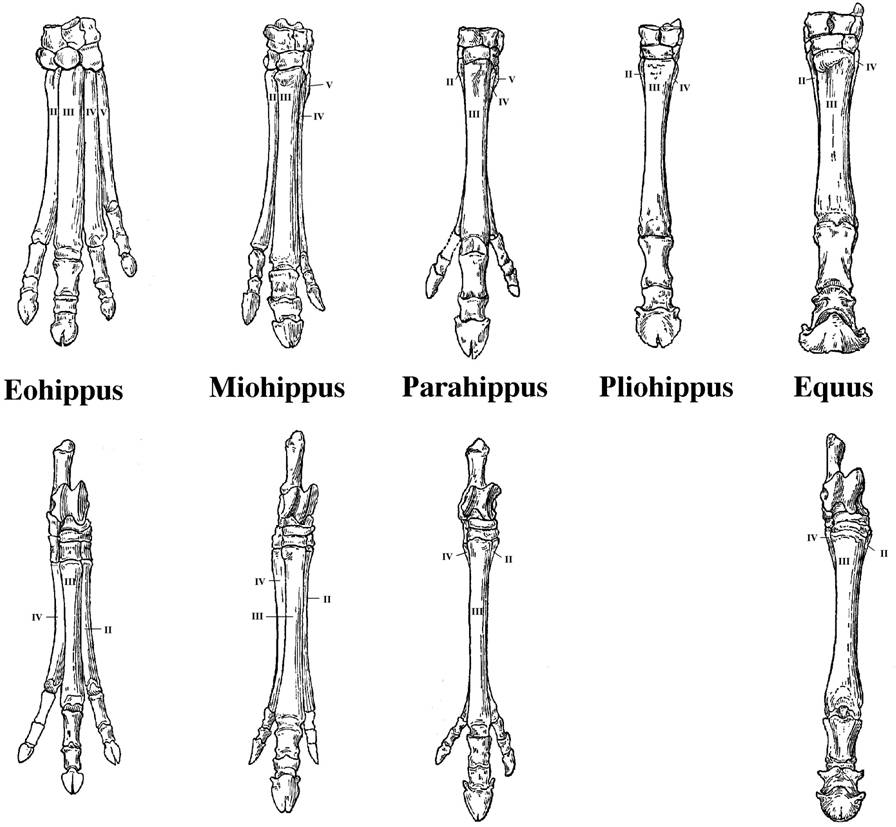
2. A second major pattern occurred
within some lineages of similar organisms. Within some lineages, we seen orderly
change in the size or characteristics of species in a geological sequence. For
instance, consider the morphological patterns in a particular taxon (horses).
Fossils in a stratigraphic sequence are similar, but often have traits that
form a continuum...like the progressive loss of digits on the horse limb. And,
with each innovation, there are often radiations - a "spurt" in the
number of species that show this new trait. And finally, these species in recent
strat are more similar to living ('extant') species than the species found in
deeper, older strata. So, many of these transitional sequences terminate in
living representatives.

 c.
Comparative Anatomy shows patterns realted to the environment:
c.
Comparative Anatomy shows patterns realted to the environment:
Homologous Structures
Although having a different outward "look" and although used
for different purposes, they have an underlying similarity in structure - forelimbs
of vertebrates all have one long upper arm bone, two lower arm bones, a bunch
of wrist bones, and five digits. Darwin saw the similarity in structure as important.
An engineer builds different things for different purposes - cars, boats, and
airplanes are structurally DIFFERENT. Here, however, it seemed as if one basic
structure was modified for different uses. Darwin knew why siblings in a family
were similar - they had the same parents (ancestors). He reasoned that these
structural similarities in different species might be due to the same principle
- common ancestry. Also, he observed a correlation: Different uses correlated
with different environments. Could this correlation be causal?
Analogous
Structures
Organisms in the same environment often have a similar outward
structure or body plan. For example, flying animals all have an aerodynamic
wing that is wider at the front than at the rear. However, the wings of differnt
animals are differnt in underlying structure. Bats have fingers that support
the membraneous wing, whereas birds lack fingers and the body of the wing consists
of feathers. Insect wings don't involve the limbs at all (even though they have
6!). Again, Darwin observed this correlation with the environment: similar use
(and outward structure) in similar environments. Could this correlation be causal?
Vestigial Organs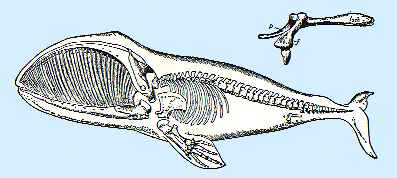
These are organs that have no function in one organism (where
they are 'vestigial') but they do function in other organisms. So, some whales
have hip bones, but no legs. Why do they have these bones? Darwin was struck
by the IMPERFECTIONS in nature, as much as the adaptations. Why do men have
nipples? Why do we have muscles that wiggle our ears? Why do we have strong
muscles in the front of our stomach, which are not "load-bearing", and weak
muscles at the base of our abdomen (which rupture in a hernia)? This is a reasonable
relationship in a quadraped, but not in a biped. Why do we have tail bones,
but no external tail? Again, these are NOT well-designed features. In fact,
attributing these imperfect designs to a perfect creator could be interpreted
as heretical. However, when we see them working in OTHER species, it suggests
that maybe we inherited them from common ancestors where they DID serve a function.
As a scientist, Darwin was trying to explain ALL the data (adaptations and imperfections),
he was not simply bringing forward only the data that supported a preferred
position (design).
d. Species distributions show patterns realtd to the environment:
Under similar environmental conditions, we find different
species filling similar ecological niches. Outward 'form' correlates with
ecological niche (role) across entire communities. So, in Australia, marsupials
fill the role of dog-like predator, cat-like predator, burrowing animal, ant-eater,
etc. These same roles are filled by outwardly similar placental mammals in South
America. However, the similarity between a wolf (placental) and a Thylacine
(marsupial - the 'tasmanian wolf') are strictly ANALOGIES. Their underlying
structure shows them to be quite different - a wolf is more similar to a ground
hog (both placentals) in underlying structure than to a thylacine. 
Islands often
have fewer species than a mainland - even a patch of mainland the same size.
As such, the patterns and interactions are often simpler to describe and understand.
For both Darwin and Alfred Russel Wallace (the other independent author of the
theory of evolution by natural selection), the study of islands was critical
in to the development of their ideas.
1. Distance correlates with the
uniqueness of the inhabitants: the animals on the Fauklands are the same
species as on the mainland, but the Galapagos fauna is composed of unique species,
found nowhere else:
"The natural history of these islands is eminently curious, and well deserves
attention. Most of the organic productions are aboriginal creations, found nowhere
else; there is even a difference between the inhabitants of the different islands;
yet all show a marked relationship with those of America, though separated from
that continent by an open space of ocean, between 500 and 600 miles in width.
The archipelago is a little world within itself, or rather a satellite attached
to America, whence it has derived a few stray colonists, and has received the
general character of its indigenous productions. Considering the small size
of the islands, we feel the more astonished at the number of their aboriginal
beings, and at their confined range. Seeing every height crowned with its crater,
and the boundaries of most of the lava- streams still distinct, we are led to
believe that within a period geologically recent the unbroken ocean was here
spread out. Hence, both in space and time, we seem to be brought somewhat near
to that great fact -- that mystery of mysteries -- the first appearance of new
beings on this earth." The Voyage of the Beagle - Darwin (1839).
 2. The Galapagos fauna:
2. The Galapagos fauna:
- It was related to american fauna,
yet different: the types of animals are new world animals.... there are iguanas
like the green iguana of Central and South America, but the iguanas are different
species. So, darwin describe it as " a world within itself, or rather,
a satellite of the Americas" .... it was different, but more like the American
fauna than any other...(no chameleons, for instance, which are old world lizards...)
- It was dominated by dispersive forms. This is critical. The communities are
dominated by reptiles, birds, and marine mammals. All of these organisms could
MIGRATE to the islands from the mainland. (Terrestrial mammals don't migrate
as well as terrestrial reptiles over open ocean. Throw a reptile in cold salty
water, and: 1) its metabolism slows down (its cold), so 2) its demand for food
and water decline; and 3) its scales protect it against water loss... which
is why reptiles do well in the desert, too. Throw a mammal in cold salt water,
and it's going to have a VERY tought time: 1) the temperature gradient between
its warm body and the cold ocean is very large - in order to maintain its high
body temperature against this gradient, it's metabolism has to INCREASE (to
produce more heat to compensate for the heat lost to the environment). This
increased metabolic demand will INCREASE the need for food and water... that's
probably in pretty short supply in the open ocean; and 2) water is lost quickly
from the skin to the salty ocean once the fur is wet... so, mammals are more
likely to starve or die of exposure than reptiles.
-
So, the islands are dominated by dispersive forms, and this suggests they came
from America. But if they came from America, WHY ARE THEY DIFFERENT SPECIES
THAN THOSE IN AMERICA? They must have changed since their arrival.
- There are even differences between
species on different islands. On the 14 species of finches - "Seeing this gradation
and diversity of structure in one small, intimately related group of birds,
one might really fancy that from an original paucity of birds in this archipelago,
one species had been taken and modified for different ends." The Voyage
of the Beagle - Darwin (1839) VIDEO
2. Argument
For Evolution as a Historical Fact:
Premise 1: Species
that are alive today are different from those that have lived previously.
Premise 2: Spontaneous Generation is refuted, so organisms
only come from other organisms.
Conclusion 1: Thus, the organisms alive today must have come
from those pre-existing, yet different, species.
Conclusion 2: There must have been change through time (evolution).
Conclusion 3: The fossil record, vestigial organs, and homologies
are all suggestive of descent from common ancestors.
Below, the figure from The Origin
of Species that shows Darwin's idea of descent from common ancestors.
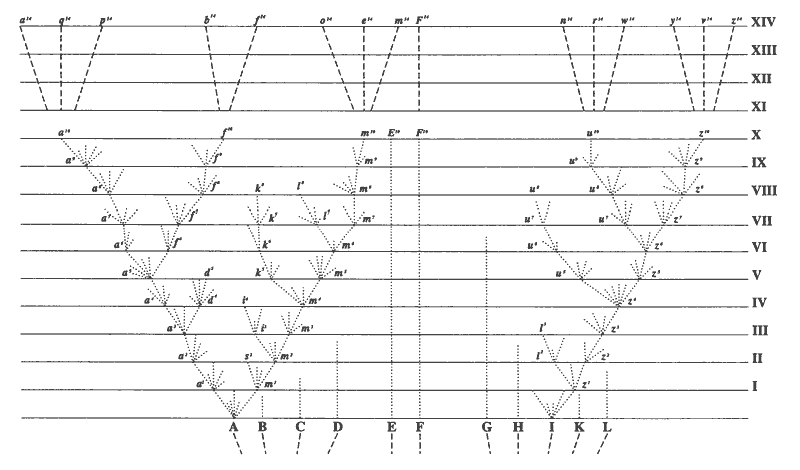
So, if species do change over time
(evolve), the next question is "How?" How does this change occur?
3. How does change occur? Natural Selection
(Know this. Understand it. You WILL be asked to outline NS in this
very form.)
P1: Populations over-reproduce (Malthus)
P2: resources are finite (Malthus)
C1: Eventually, a population will grow until it becomes limited by its resources.
At that time, their will be a "struggle for existence" and most offspring
produced will die. (Malthus)
P3: Individuals in a population vary, and some of this variation is heritable (Darwin
- observations and animal/plant breeding)
C2: Variations will not have the same probability of survival and reproduction in
a particular environment; those well-suited to the environment will be more
likely to survive and reproduce than others, passing on the genes for these
adapted traits. There will be "Differential Reproductive Success" (Observations,
breeding).
C3: Over time, adaptive traits will accumulate and the characteristics in a population
will change. This is lineage evolution. (Like change in horse toes in a sequence
of fossil species, or like the change in the chihuahua lineage from the ancestral
wolves).
Corollary: Two sub-populations, separated in different environments, would be selected
for different traits and may subsequently lose the capacity to interbreed. At
this point, they are different biological species. This is Speciation and Radiational
Evolution. (like the production of different Finches, mockingbirds, etc. on
different islands in the galapagos, and like the radiation of St. Bernards AND
chihuahua's, which diverged from one another over time).
Darwin here provides a natural explanation
for why purposeful structures and behaviors occurs in nature. Through some process
unknown to him, variation arises in natural populations. These varieties differ
in terms of functional efficiency in a common environment; so some improving
an organisms probability of surviving and mating than others. Organisms with
these beneficial traits will leave more offspring, and the frequencies of these
beneficial characteristics will increase through time - much as humans select
for smaller and smaller dogs. He ends The Origin of Species (1859)
like this:
 "It
is interesting to contemplate an entangled bank, clothed with many plants of
many kinds, with birds singing on the bushes, with various insects flitting
about, and with worms crawling through the damp earth, and to reflect that these
elaborately constructed forms, so different from each other, and dependent on
each other in so complex a manner, have all been produced by laws acting around
us. These laws, taken in the largest sense, being Growth with Reproduction;
Inheritance which is almost implied by reproduction; Variability from the indirect
and direct action of the external conditions of life, and from use and disuse;
a Ratio of Increase so high as to lead to a Struggle for Life, and as a consequence
to Natural Selection, entailing Divergence of Character and the Extinction of
less-improved forms. Thus, from the war of nature, from famine and death, the
most exalted object which we are capable of conceiving, namely, the production
of the higher animals, directly follows. There is grandeur in this view of life,
with its several powers, having been originally breathed into a few forms or
into one; and that, whilst this planet has gone cycling on according to the
fixed law of gravity, from so simple a beginning endless forms most beautiful
and most wonderful have been, and are being, evolved". - The Origin of Species (Darwin 1859).
"It
is interesting to contemplate an entangled bank, clothed with many plants of
many kinds, with birds singing on the bushes, with various insects flitting
about, and with worms crawling through the damp earth, and to reflect that these
elaborately constructed forms, so different from each other, and dependent on
each other in so complex a manner, have all been produced by laws acting around
us. These laws, taken in the largest sense, being Growth with Reproduction;
Inheritance which is almost implied by reproduction; Variability from the indirect
and direct action of the external conditions of life, and from use and disuse;
a Ratio of Increase so high as to lead to a Struggle for Life, and as a consequence
to Natural Selection, entailing Divergence of Character and the Extinction of
less-improved forms. Thus, from the war of nature, from famine and death, the
most exalted object which we are capable of conceiving, namely, the production
of the higher animals, directly follows. There is grandeur in this view of life,
with its several powers, having been originally breathed into a few forms or
into one; and that, whilst this planet has gone cycling on according to the
fixed law of gravity, from so simple a beginning endless forms most beautiful
and most wonderful have been, and are being, evolved". - The Origin of Species (Darwin 1859).
II. Testing Darwin's Theories
A. The Fossil
Evidence
1. Radioactive
Decay and Geological Clocks
Darwin had guessed that the earth
had to be at least 300 million years old to explain the evolution of life through
the stately process of natural selection. W. Thompson ("Lord Kelvin")
was a physicist who demonstrated that the Earth could be no more than 24 million
years old. He did this by calculating how long it would take for a molten
object with the Earth's mass to cool to the current temperature of the Earth.
However, he made his predictions before Becquerel and the Curies discovered
radioactivity - a process that releases heat and keeps the Earth warmer than
it "should be" (based on Thompson's inference). After the Curies discovered
radium and studied radiactivity (1898), Ernst Rutherford realized that the regular
rate of decay could be used to age rocks.
a.
Principle:
- measure amt of parent and daughter isotopes = total initial parental
- with the measureable1/2 life, determine time needed to decay this fraction
- K40-Ar40 suppose 1/2 of total is Ar40 = 1.3by
(Now, you might say "be real"! How
can we measure something that is this slow?)
Well, 40 grams of Potassium (K)
contains:
6.0 x 1023 atoms (Avogadro's number, remember that little chemistry
tid-bit?). So, For 1/2 of them to change, that would be:
3.0 x 1023 atoms in 1.3 billion years (1.3 x 109)
So, divide 3.0 x 1023 by 1.3 x 109 = 2.3 X 1014 atoms/year.
Then, divide 2.3 x 1014 by 365 (3.65 x 102) days per year
= 0.62 x 1012 atoms per day ( shift decimal = 6.2 x 1011)
Then, divide 6.2 x 1011 by 24*60*60 = 86,400 seconds/day: (= 8.64
x 104) = 0.7 x 107 atoms per second
0.7 x 107 = 7 x 106 = 7 million atoms changing from Potassium
to Argon every second!!!
This 'decay' gives off energy - radiation
that is detectible and measureable by Geiger counters and similar instruments.
It is actually an easily measured rate. And you can come back and measure
it again tomorrow, next week, next year, or in 50 years; and it has always been
the same.
b. Different
Clocks
The K-Ar clock is ok for measuring
things that happened billions of years ago, but it is not a good clock for things
happening hundreds or thousands of years ago because the rate of change is so
slow. It would be like measuring an olympic 100m sprint (which last less than
10 seconds these days) with a 2-minute hour-glass egg timer. After ten seconds,
only a few grains of sand will have fallen. For measuring anything, you need
a gauge with the correct level of resolution. For an olympic 100m sprint, an
electronic 'stopwatch' measuring to the thousandths of a second is needed. For
measuring the marathon, you don't want that type of resolution; an analog clock
that measures hours, minutes, and seconds is probably appropriate. Of course,
when you measure the same event with different types of clocks, they should
all give roughly the same answer, even if the answers differ in precision. Luckily,
there are different isotopic series that give resolution at different time scales:
C14 - decays to C12 with a half-life
of 5730 years
Longer periods - use different elements with longer half-lives
- K40 decays to Ar40 at a rate of 50%/1.3 b years
- U238 - Pb208 = 4.5 by
- Rb87 - St87 = 47 by
- U238 - Thorium 230 = 80,000
- U235 - Protactinium231 = 34,000
c. Tests
of Corroboration
We gain greater confidence in a conclusion
if it is supported by multiple, independent pieces of evidence. One eyewitness
to a crime is ok, but three different people who all tell the same story are
far more convincing. Because the decay process in one element
present in rock has no effect on the way other elements decay, the decay series
are independent of one another. So, one rock could be aged using the K-Ar clock,
the Rb-St clock, and the U-Pb clock. If we get roughly the same age using these
three different clocks, we would be more confident in that age. There are other
corroborating methods, too, that don't involve decay, at all. For example, we
can carbon-date material from Pompeii, which we know from historical record
should date to 79ACE. Indeed, they do, corroborating the use of radiocarbon
dating. One of the most dramatic corroborations involves predictions from astronomers.
Astronomers have postulated that, because of tidal friction between the water
in the ocean and the earth, the rotation of the Earth has been slowing down
over time. In fact, astronomers have calculated that, based on the masses of
the oceans and the Earth, and the frictional coefficient of water against land,
that the length of an Earth's rotation (a day) has slowed at a rate of 2 seconds
every 100,000 years. (So days were SHORTER in the past and have SLOWED to the
current 24 period). This process would not affect the time it takes the
Earth to orbit the sun, however. So, it would take a year for the Earth to orbit
the sun, but the Earth would be spinning on its axis more rapidly in the past.
This means that a year in the past would have MORE days than it does now.
Indeed, astrophysicists predict that, based on tidal friction, a year 380 million
years ago would have contained 400 days. Wow... crazy. Well, corals lay down
a layer of calcium carbonate 'shell' every day. There are seasonal changes
in the thickness of the layers, so years can be distinguished, too. Some
corals have been dated by radioactive decay to be 380 million years old. If
this age based on decay is correct, and if the astrophysicists are correct,
then they should have 400 daily layers of growth within each yearly band.
They do. So, we can use basic physics to predict the "age" of a coral.
And when we compare that "age" to the "age" determined by radioactive decay,
the ages are the same. So, either they are both true, or they are both wrong
in the same way. It is unlikely that we have such basic physics wrong.
Very unlikely. So, the Earth is, indeed, very very old. Radioactive decay
is constant; if it wasn't, or if it hadn't been in the past, none of these comparisons
would work. period. But they DO work, and so it is irrational to
conclude that the Earth is young, or that radioactive dating doesn't work or
is somehow "dubious". Again, our ability to harness the power of radioactive
decay in nuclear reactors is powerful testimony to the degree of confidence
we have in our knowledge and understanding of the decay process. And thus, we
have great confidence in the great age of the Earth.
2.
'Transitional' fossils
One of Darwin's dilemmas was the
lack of continuous sequences of fossils that preserve a complete record of evolutionary
change. In 1859, the fossil record was best described as 'discontinuous' for
most lineages. Of particular interest to Darwin's model of common descent was
the absence of 'transitional' fossils - fossils that showed the nascent beginnings
of a major new type of organism evolving from more primitive stock. In the last
150 years, paleontology has unearthed millions of fossils and many have been
placed in very complete sequences. They provide a solution to Darwin's dilemma
and also allow us to reconstruct phylogenies. We will take a look at some of
the more remarkable transitional sequences that have been documented in the
evolution of vertebrates.
Transitional fossils are important
in two ways. First, they contain
a complement of traits that makes them hard to pigeonhole into one group of
organisms or another. In other words, they have a combination of traits from
two separate groups (an intermediate morphology).
But this is not all. I mean,
there are alot of crazy organisms out there. The existence of a weird
combination of traits does not mean the organism is necessarily a transitional
form, nor does this support common ancestry in and of itself. Evolution does
something more than simply predict the existence of transitional forms - if
predicts WHEN these forms should exist. Let's apply these ideas to some real
fossils.
a.
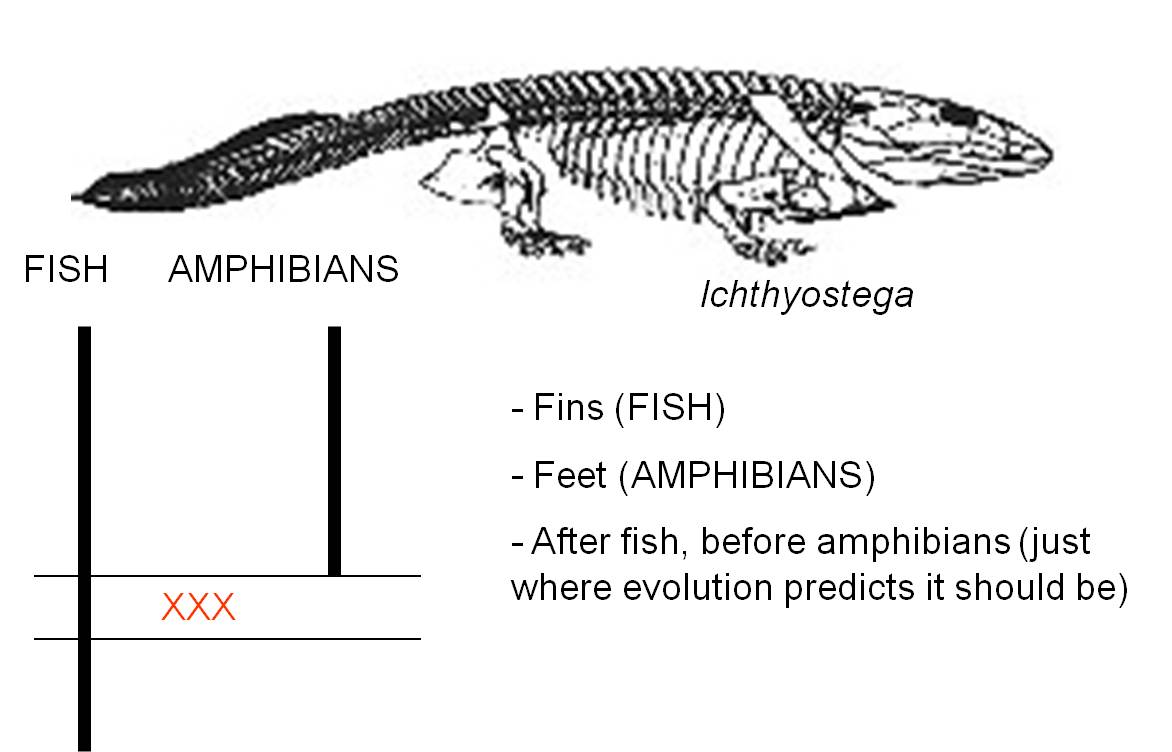
Ichthyostega spp. and
the evolution of tetrapods
Darwin hypothesized that amphibians
evolved from fish. This created a few important problems - how did lungs and
feet evolve? In 1929, several species with a mix of fish and amphibain characters
were discovered in Greenland and placed within the genus Ichthyostega.
These animals had lungs and gills - an obviously intermediate morphology. And
they had tails with cartilagineous rays in the fin, like a fish tail. But these
animals also had feet. The legs were probably not strong enough to bear the
entire weight of the animal, but it was a true tetrapod. So, it had a suite
of fish and amphibian traits. However, that is not enough. To be a transitional
fossil and to truly test the hypothesis that this animal is an ancestor of amphibians
and a descendant of fish, it must come after true fish and before true amphibians.
Indeed, that is right where it is.
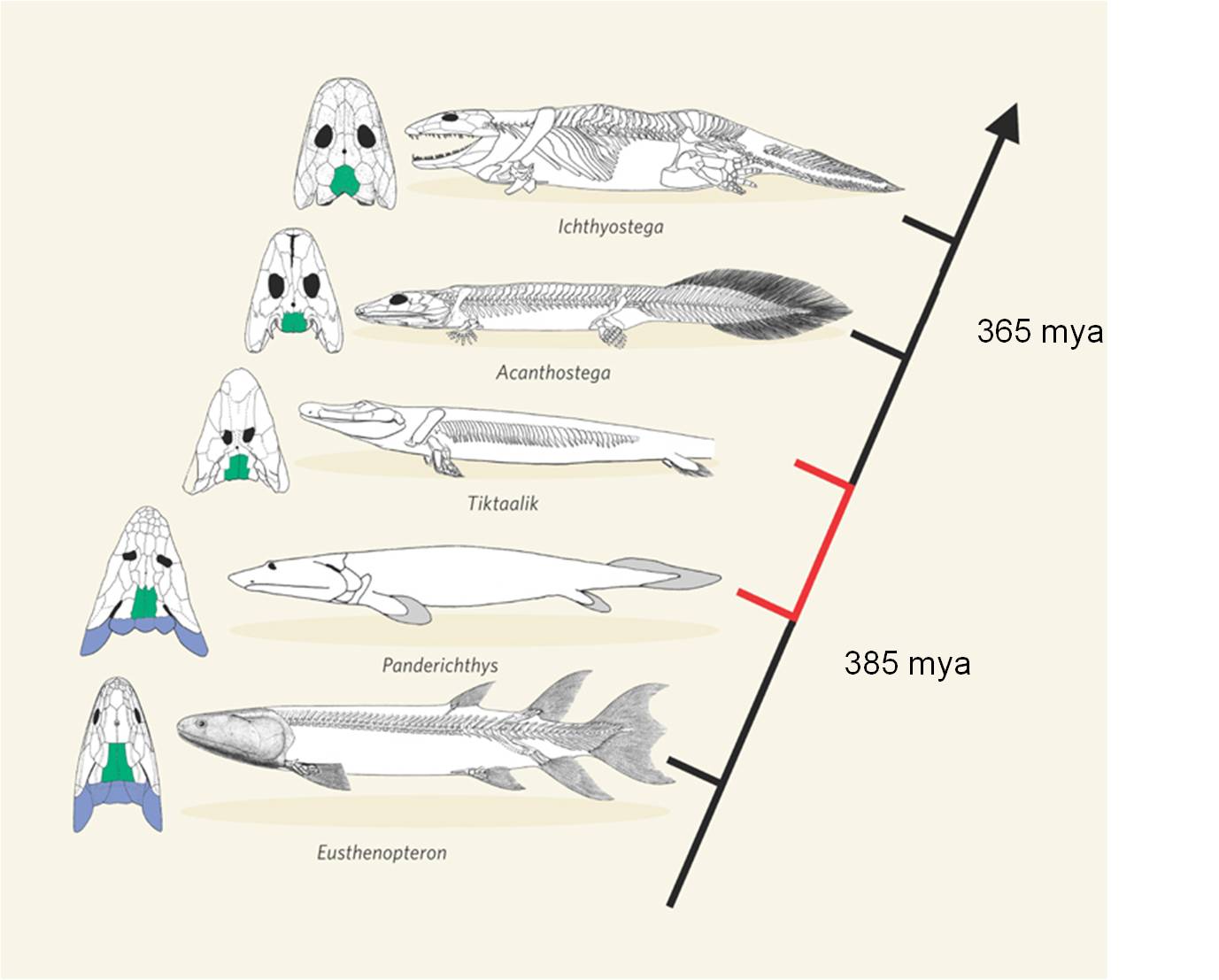
This transition between fish and
land animals was one of the most important evolutionary transitions in the history
of life. Since 1929, many fossils have been found that form a very complete
transitional sequence, linking lobe-finned fish like Panderichthyes
to proto-amphibians like Ichthyostega and Acanthostega. In
2004, an exceptional fossil was found in Northern Canada that was intermediate
between Panderichthyes and Ichthyostega. Tiktaalik rosae
was such a beautiful transitional form that it was dubbed the "fish-o-pod".
The animal has short bony forelimbs that terminate in fins, not feet. However,
the limbs are able to support weight, and have a wrist-like joint that allows
the limb to pivot and propel the animal forward. It has a decidedly fish-like
body, but an Ichthyostegan head and intermediate limbs.
 |
 |
| Ichthyostega was one of the first fossils that bridged
the gap between fish and amphibians |
Now, a series of intermediates shows the transition from ancestral, lobe-finned
fishes, through limbed fishes like the "fish-a-pod" Tiktaalik,
to amphibians |
b.
Archeopteryx lithographica
and the evolution of birds
Archeopteryx has an intermediate
morphology containing reptilian and avian characteristics. It has fingers, teeth,
and a bony tail like reptiles (and unlike birds), but it has feathers like birds
(and unlike modern reptiles). So, it had a combination of traits from two major
groups. No birds today have teeth or fingers, and no reptiles have feathers.
So, it is intermediate in morphology. But evolution PREDICTS something
else about this organism. IF it was a biological link between reptiles
and birds, then it would have to have lived after other reptiles (who
were its ancestors) and BEFORE all true birds (who might be its descendants).
It is. The hypothesis has
been tested by evidence from the physical world. Evolution is a testable, supported
theory. Indeed, since 1861, further paleontological evidence suggests that birds
didn't evolve from just any reptile, but from a specific group called the Maniraptoriformes.
This group diverged from the Tyrannosauroidea, which includes Tyrannosaurus
rex. Within the maniraptoriformes, we see a wide variety of feathered reptiles
discoverd in China since 1990. However, the feathers are not only on the limbs
and they are definitely not used for flight. In fact, in the oldest fossils
of feathered dinosaurs, the protofeathers are only on the head and spine. A
likely scenario is that feathers were first used for attracting mates. Selection
for increased feather distribution provided the additional benefit of insulation
and homeothermy. Finally, large feathers on the limbs might provide lift while
climbing, running, or gliding, and selection could favor the acquisition of
flight. This provides a nice example of how a complex trait - flight feathers
- might NOT have evolved for that purpose intially.
 |
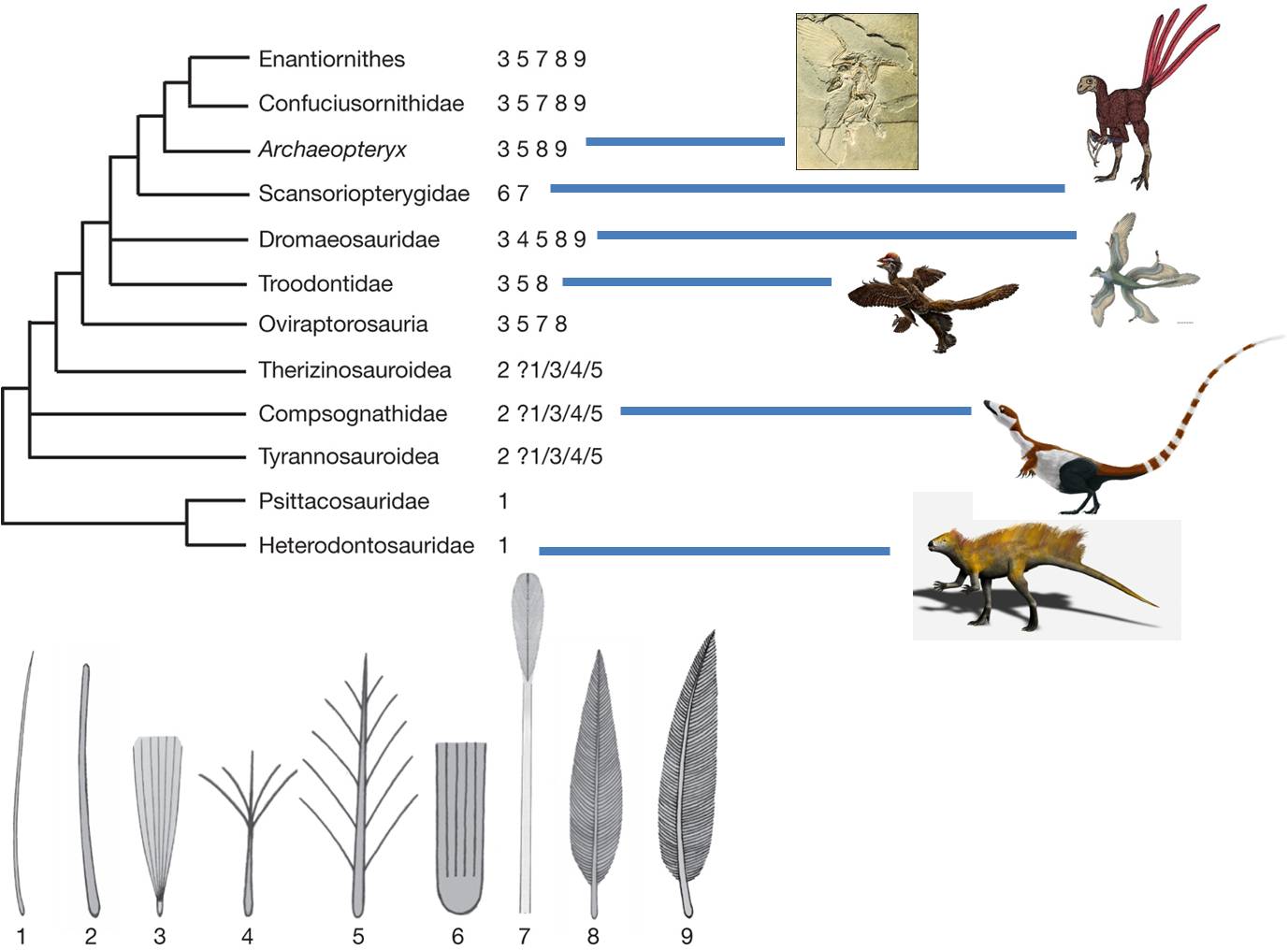 |
| Archaeopteryx was the first intermediate discovered
between reptiles and birds. |
Now, paleonotologists have found numberous lineages of 'feathered dinosaurs',
showing that the lineage leading to Archaeopteryx and modern birds
was only one branch of ancestral, feathered animals. |
c. Therapsids
and the evolution of mammals
The transition from reptiles to mammals
is one of the most well-document transitions in the fossil record. Indeed, there
is such a nice sequence that it is difficult to specify where the most important
or instructive transition occurs. For our purposes, we will look at a group
of organisms called the therapsids. Like reptiles, therapsids have several bones
in their lower jaw, and one inner ear bone. Like mammals, they had specialized
detition with incisor-like teeth at the front,and larger canines. Also like
mammals, they walked with their legs underneath them, rather than out to the
side like ancestral reptiles. Through the therapsid lineages, we have a very
clear sequence of transitions that show how several of the lower jaw bones of
reptiles became reduced and were eventually used as inner ear bones in the mammals.
And of course, the therapsids fill the temporal gap between one group of ancestral
reptiles and the more modern mammals - just as evolutionary theory and common
ancestry predict.
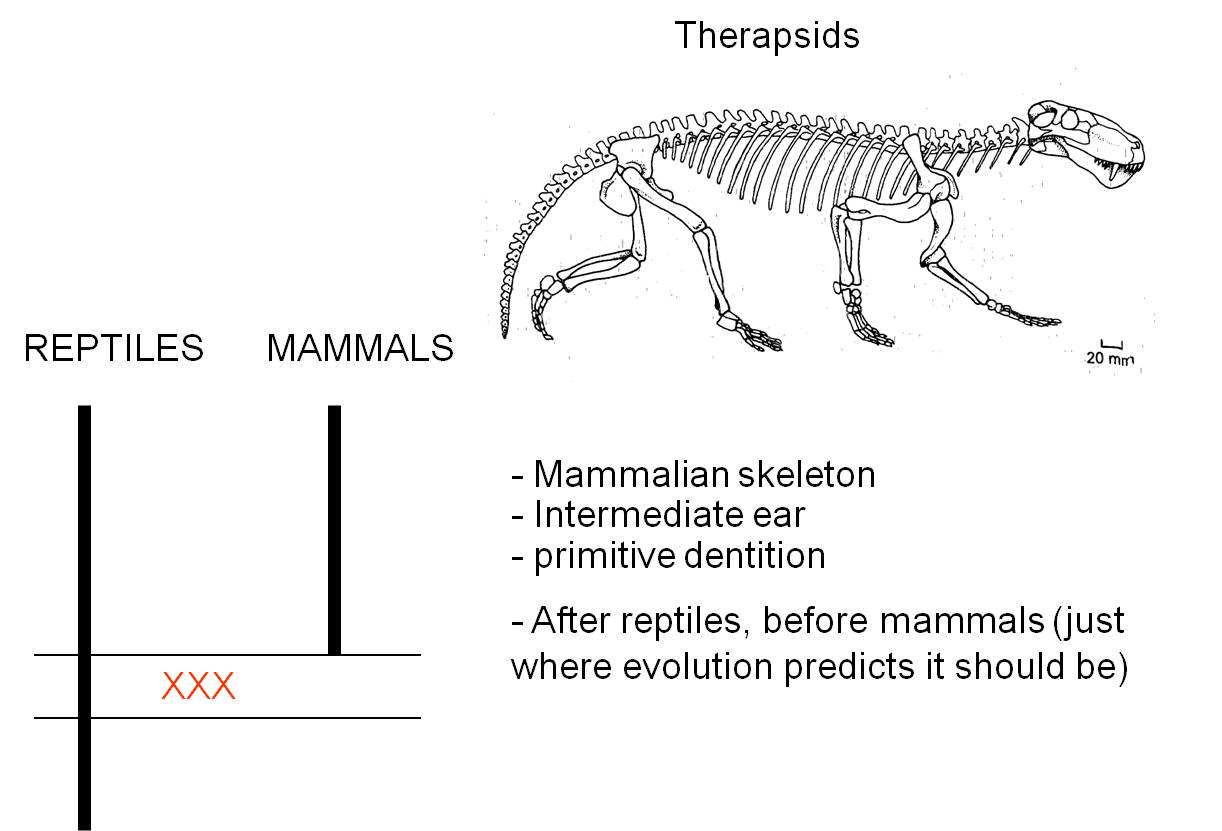 |
 |
| Therapsids were a group of reptiles that dominated during the Permian
Period, 250 million years ago. |
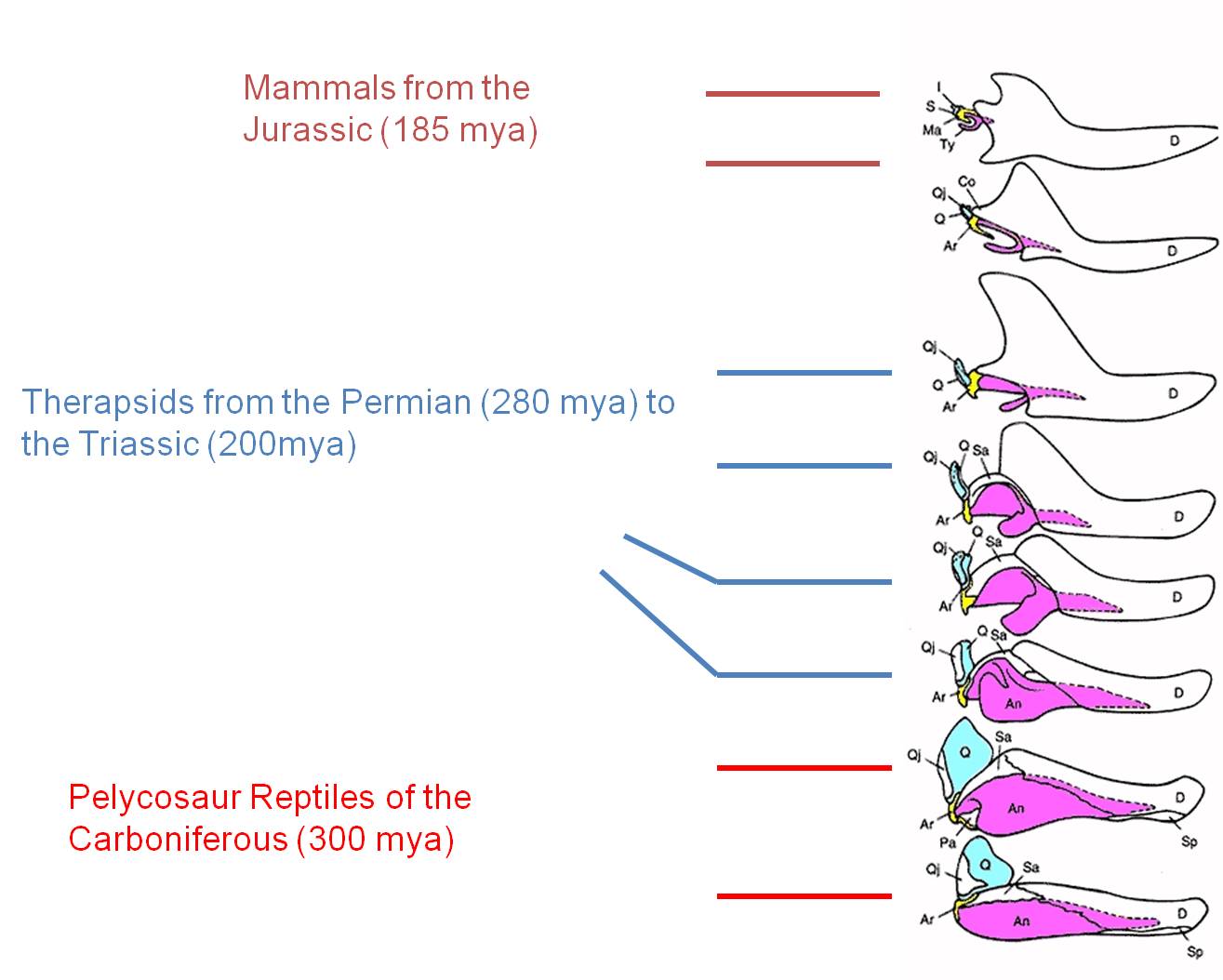
The evolution of the mammalian middle ear bones is beautifully preserved
in the fossil record. The three middle ear bones in mammals (blue, yellow,
pink) are homologous and descended from lower jaw bones of reptiles that
became reduced in size and took on another function as their role in reptilian
jaw function was assumed by the dentary bone (white). |
d. Australopithecines
and the evolution of humans
Even Linnaeus recognized the morphological
similarity between humans and apes (chimpanzees, gorillas, and orangutans).
When we look at these species, the things that set humans apart are our upright
stance and bipedal locomotion, our large heads, and our relatively short forelimbs.
Ever since the first neanderthal fossil was found in 1856 and after Darwin wrote
The Descent of Man in 1871, we have been reconstructing the phylogeny
of our own species. There are many fossil species that form an excellent transitional
sequence between ancestral primates and modern humans, again making it difficult
to pick just one group. However, The Australopithecines provide a good and historically
important group. In 1924, Raymond Dart discovered a very small skull of a juvenile
primate that he named Australopithecus africanus, meaning "southern
man-ape from Africa". Like other australopithecine fossils to follow, it
had a short but ape-like snout with reduced canines. Dart suggested that the
presence of this fossil in Africa, and the presence of chimps and gorillas in
Africa, confirmed Darwin's hypothesis of an African origin of humans. This was
highly debated by other anthropologists who believed that humans evolved in
Europe or Asia. The discoveries of the Leakey's in Olduvai Gorge in the 1950's
and 1960's, and the discovery of Australopithecus afarensis in the
1970's by Donald Johanson, supported the African origins model. The most complete
single fossil of A. afarensis, known as the 'Lucy" fossil, shows
the combination of traits expected in an intermediate, transitional species.
The hip and the articulation of the femur and tibia show that the organism walked
erect - a characteristic that is distinctly human. However, the cranial volume
is very small - only 25% the volume of modern humans and equal to the volume
of chimpanzees. However, other facial features are intermediate; the snout is
shorter than in chimps and gorillas, but the canines are much larger than in
humans. And again, these fossils fall before more human species and after more
primitive primates; just as common ancestry and evolution would predict.
Yes, this is redundant; but it is
redundant for a reason. There aren't just one or two fossils that 'conform'
to the expectations of evolutionary theory. There are 100's of intermediates
that provide tests and confirmation of evolutionary theory. One of the most
frequent claims of creationists is that "there are no intermediate fossils".
Well, you've seen quite a few, linking the major types of vertebrates.
Darwin's theory of common ancestry predicted their existence, and scientists
have tested this prediction by looking for physical evidence that could test
this hypothesis. Although only Archeopteryx and neanderthals were
discovered in his lifetime, we now have intermediates linking all major groups
of vertebrates, confirming these hypotheses. It makes you wonder why these
claims continue to be made, in the face of such overwhelming physical evidence.
 |
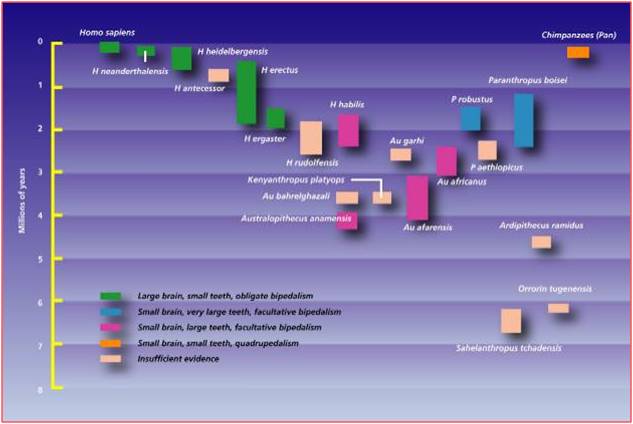 |
| When discovered in 1974, Australopithecus afarensis was the oldest
fossil of a bipedal hominid. |
Since then, several more primitive bipedal species have been discovered.
The fossil history of the hominid lineage has been very well described. |
C. Genetic
Evidence
 1.
Gross Chromosomal Similarities
1.
Gross Chromosomal Similarities
The most definitive tests of biological
relatedness come by examining DNA. Why? Because the only place an organism
gets its DNA is from its parents, their parents, their great-grand parents,
and their ancestors. Barring the rare events of lateral gene transfer that can
occur in some organisms, the only reason two organisms would have similar DNA
is that they are biologically related. So, If I am accused of fathering a child,
and my DNA is similar to that child's DNA, then I can be "convicted" of being
related to that child. That is the only reason two organisms will share
DNA - because they are biologically related. This pattern is reinforced
by our understanding of meiosis and sexual reproduction, which explain why these
patterns of relatedness occur. Now, when we see similarities among species in
DNA structure, logical consistency demands that we propose the same hypothesis
for the same pattern. In the figure at right, you see the chromosomes from a
human, a chimp, a gorilla, and an orangutan. The most striking thing is the
similarity in banding patterns across these chromosomes. Remember what those
bands signify? The dark areas are heterochromatin, that have a low concentration
of coding sequences. The lighter areas are euchromatin, where most of the genes
are. So, we are looking at similarities in the large scale architecture of the
genomes from these organisms. Evolutionary theory predicted that humans would
have similar DNA to apes, and they do - even at the level of gross chromosomal
structure. However, there is a major difference here; humans have n=23 while
the other species have n=24. How can evolutionary biology explain this difference
in chromosome number? Well, even the exception here proves the rule. The long
#2 chromosome in humans - the first chromosome in thes econd set of chromosomes
in the upper left corner of the figure - is banded like two of the chromosomes
in other primates (shown next to it). A simple hypothesis would be that, at
some point in the human lineage after divergence with chimp-like ancestors,
these two chromosomes fused and became inherited as one unit. Can you remember
an instance where chromosomes get stuck together and inherited as a single unit?
It happens in translocation events, like in translocation Downs. And of course,
this is not always deleterious to the organism - carriers for the translocation
chromosome are phenotypically normal. A mating between two carriers could produce
an offspring with the correct DNA content, but with two fewer chromosomes. Apparently,
just such a modification may have occurred in the human lineage after divergence
for the common ancestor we share with chimpanzees.
2. Mutational
clocks
Mutations occur over time; the longer
populations diverge from one another, the more mutational differences should
accumulate between them. Many mutations have little effect on the phenotype
- indeed, mutations in the non-coding intron regions of a gene, or in non-coding
sequences between genes, have no effect on the phenotype. These mutations will
be selectively neutral - and they should accumulate at a steady rate over time.
If we can measure the mutation rate, then we can use this rate like a 'clock':
we can count the number of mutational differences there are between the DNA
from differnt populations, and then compute how long they must have diverged
from one another to account for the genetic difference that we see.
3. Genetic
Phylogenies
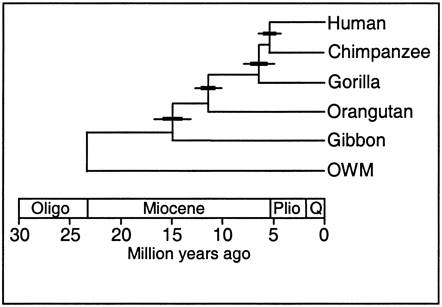 Because
DNA comes from ancestors, similarity in DNA implies common ancestry. The greater
the similarity, the less time since their common ancestor; in other words, the
more "recent" their common ancestry. For example, human DNA and chimp
DNA is 98.4% similar in nucleotide
sequence, so they share a more recent common ancestor than either species shares
with gorillas - which are similar to both humans and chimps at a rate of 95%.
Now, some might claim that these similarities are analogous, representing
similarities between organisms that function in a similar way (but are not biologically
related). But, only 10% of the genome is a recipe for protein. Even
the 90% that does not code for protein, that is random sequence, still shows
this similarity. Even non-functional DNA is similar, so functional similarity
(ie., ANALOGY) can't be the answer... the similarity must be HOMOLOGOUS - the
result of common ancestry. Genetic phylogenies have been a powerful tool for
reconstructing the evolutionary relationships among broad categories of organisms
that are very different or similar morphologically. So, for instance, biologists
had long believed that all prokaryotes were closely related to one another,
and not closely related to the eukaryotes. Genetic analyses revealed, however,
that the Archaeans were more closely related to eukaryotes than to the other
prokaryotic group, the eubacteria. Likewise, genetic analyses show that fungi
are more similar to animals than they are to plants, and green algae is more
similar to plants than they are to other forms of algae. These relationships
can be understood in an evolutionary context. The eukaryotes evolved from a
type of prokaryote - and so should be more similar genetically, to this parental
stock of prokaryotes (the archaeans) than other prokaryotes (eubacteria). Green
algae gave rise to terrestrial plants, and so should be more similar, genetically,
to this closely related group (plants) than to other algal groups. This new,
more realistic view of the history of life is reflected in a new way to classify
organisms - based on common ancestry rather than common morphology, alone. For
example, as described above, some reptile groups are more similar to birds than
to other reptiles, and some reptile groups (though extinct) are more similar
to mammals than other reptiles. The new classification scheme takes a more systematic
approach, and groups organisms based on their phylogenetic relationships (grouping
crocodilians with their close relatives the birds, for instance), rather than
grouping organisms based on shared primitive characteristics (grouping crocodiles
with turtles, snakes, and lizards in "the reptilia" because they have
scales and lay shelled eggs). We will take a look at this later in the course.
Because
DNA comes from ancestors, similarity in DNA implies common ancestry. The greater
the similarity, the less time since their common ancestor; in other words, the
more "recent" their common ancestry. For example, human DNA and chimp
DNA is 98.4% similar in nucleotide
sequence, so they share a more recent common ancestor than either species shares
with gorillas - which are similar to both humans and chimps at a rate of 95%.
Now, some might claim that these similarities are analogous, representing
similarities between organisms that function in a similar way (but are not biologically
related). But, only 10% of the genome is a recipe for protein. Even
the 90% that does not code for protein, that is random sequence, still shows
this similarity. Even non-functional DNA is similar, so functional similarity
(ie., ANALOGY) can't be the answer... the similarity must be HOMOLOGOUS - the
result of common ancestry. Genetic phylogenies have been a powerful tool for
reconstructing the evolutionary relationships among broad categories of organisms
that are very different or similar morphologically. So, for instance, biologists
had long believed that all prokaryotes were closely related to one another,
and not closely related to the eukaryotes. Genetic analyses revealed, however,
that the Archaeans were more closely related to eukaryotes than to the other
prokaryotic group, the eubacteria. Likewise, genetic analyses show that fungi
are more similar to animals than they are to plants, and green algae is more
similar to plants than they are to other forms of algae. These relationships
can be understood in an evolutionary context. The eukaryotes evolved from a
type of prokaryote - and so should be more similar genetically, to this parental
stock of prokaryotes (the archaeans) than other prokaryotes (eubacteria). Green
algae gave rise to terrestrial plants, and so should be more similar, genetically,
to this closely related group (plants) than to other algal groups. This new,
more realistic view of the history of life is reflected in a new way to classify
organisms - based on common ancestry rather than common morphology, alone. For
example, as described above, some reptile groups are more similar to birds than
to other reptiles, and some reptile groups (though extinct) are more similar
to mammals than other reptiles. The new classification scheme takes a more systematic
approach, and groups organisms based on their phylogenetic relationships (grouping
crocodilians with their close relatives the birds, for instance), rather than
grouping organisms based on shared primitive characteristics (grouping crocodiles
with turtles, snakes, and lizards in "the reptilia" because they have
scales and lay shelled eggs). We will take a look at this later in the course.
D. Concordant Phylogenies
Both the
fossil record and the pattern of genetic similarity among living species are
presented as evidence of evolution and descent from common ancestors. If BOTH
patterns due to the same phenomenon (common descent), then their patterns should
be the same. In short, there should only be one tree of life, and both
patterns should reveal that same tree. Indeed, we should be able to test the
theory of evolution yet again, in a most remarkable way: we should be able to
use the degree of genetic divergence to predict where (really "when"),
in the sedimentary strata of the earth's crust, the common ancestor of two groups
should be. Then, we should be able to go to that strata and find that common
ancestral species.
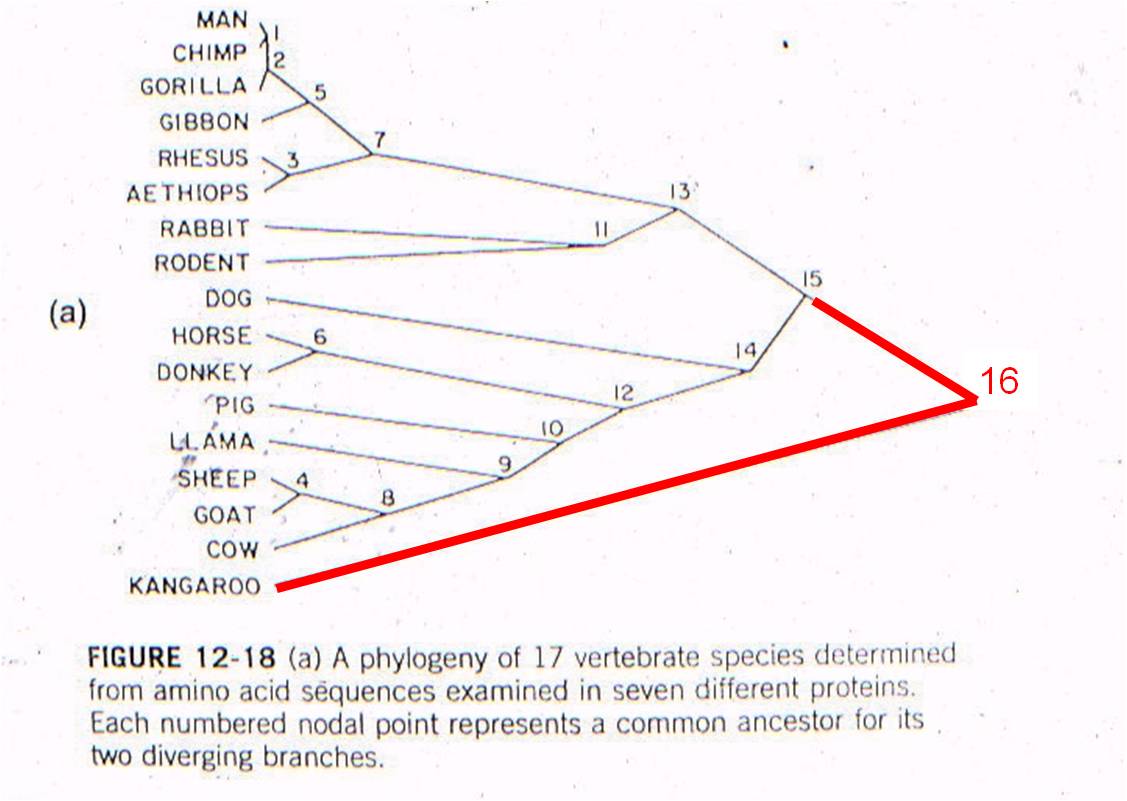
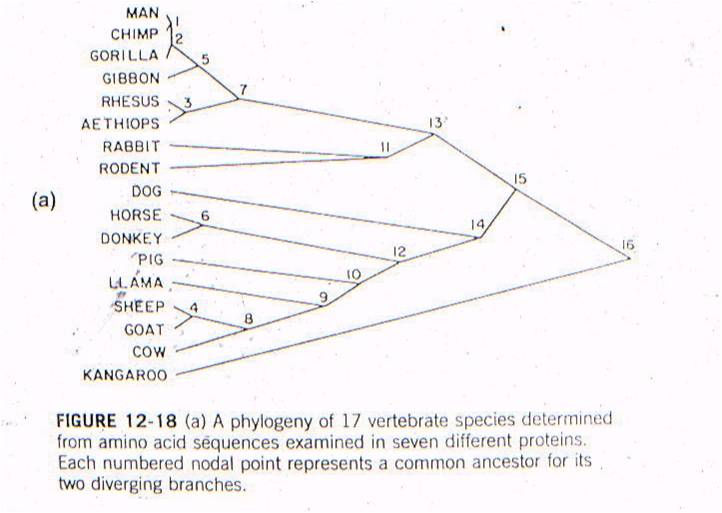 Let's
see an example of this type of test. All vertebrates have many of the same proteins,
but these proteins can differ in specific amino acid sequence. In this case,
the amino acid sequences for the same 7 proteins were sampled from 17 mammals.
For each possible pair of species, the minimum number of nucleotide substitutions
in the DNA, needed to explain the differences in the amino acid sequences, were
determined. For example, suppose humans have an argenine as the third amino
acid in collagen, while cows have lucine. Argenine is encoded by the codons
CGU, CGG, CGC, and CGA. Leucine is eoncoded by the codons CUU, CUG, CUC, and
CUA. So, although a two-base change could be responsibile (from CGU to CUA),
the minimum number of substitutions would be 1 - with just a change in the second
position (CGU to CUU). So, the minimum number of substitution mutations necessary
to explain all the sequence differerences between every pair of species is computed,
and then species are linked together based on sequence similarity. The number
at each "node" refers to the order of the clustering. So, the most similar pair
of species, of all the possible pair-wise combinations among these 17 mammals
(255 pairwise combinations) is humans and chimps - they are linked at 'node
1'. Then, gorillas are more similar to humans and chimps than any other
pair of taxa, so gorillas link to humans and chimps at node 2. Then, the
next most similar pair of taxa are Rhesus monkeys and Aethiops monkeys, linked
at node 3, and so forth. All placental mammals link together with one
another before any link to the sole marsupial, the kangaroo.
Let's
see an example of this type of test. All vertebrates have many of the same proteins,
but these proteins can differ in specific amino acid sequence. In this case,
the amino acid sequences for the same 7 proteins were sampled from 17 mammals.
For each possible pair of species, the minimum number of nucleotide substitutions
in the DNA, needed to explain the differences in the amino acid sequences, were
determined. For example, suppose humans have an argenine as the third amino
acid in collagen, while cows have lucine. Argenine is encoded by the codons
CGU, CGG, CGC, and CGA. Leucine is eoncoded by the codons CUU, CUG, CUC, and
CUA. So, although a two-base change could be responsibile (from CGU to CUA),
the minimum number of substitutions would be 1 - with just a change in the second
position (CGU to CUU). So, the minimum number of substitution mutations necessary
to explain all the sequence differerences between every pair of species is computed,
and then species are linked together based on sequence similarity. The number
at each "node" refers to the order of the clustering. So, the most similar pair
of species, of all the possible pair-wise combinations among these 17 mammals
(255 pairwise combinations) is humans and chimps - they are linked at 'node
1'. Then, gorillas are more similar to humans and chimps than any other
pair of taxa, so gorillas link to humans and chimps at node 2. Then, the
next most similar pair of taxa are Rhesus monkeys and Aethiops monkeys, linked
at node 3, and so forth. All placental mammals link together with one
another before any link to the sole marsupial, the kangaroo.
Now, this is just a clustering
procedure. It could be done on cars, nuts and bolts, anything. But
since it is done on life forms, we can test an evolutionary prediction.
Evolution suggests that organisms are similar because of common descent from
shared ancestors - represetned by these nodes. Species that are more similar,
genetically, should have a more recent ancestor than organisms that are more
different, genetically. So, there should be a relationship between 'time
since divergence' and 'genetic difference', as we explained above (mutational
clocks). Well, using the group of organisms that we have, we can describe what
evolution predicts that relationship should be:
IF: the oldest mammal in the fossil
record is ancestral to all more recent mammals, and
IF: the most different groups of mammals today (placentals and marsupials) are
descended from that ancestor, and
IF: the accumulation of genetic differences (mutations) occur at a constant
rate,
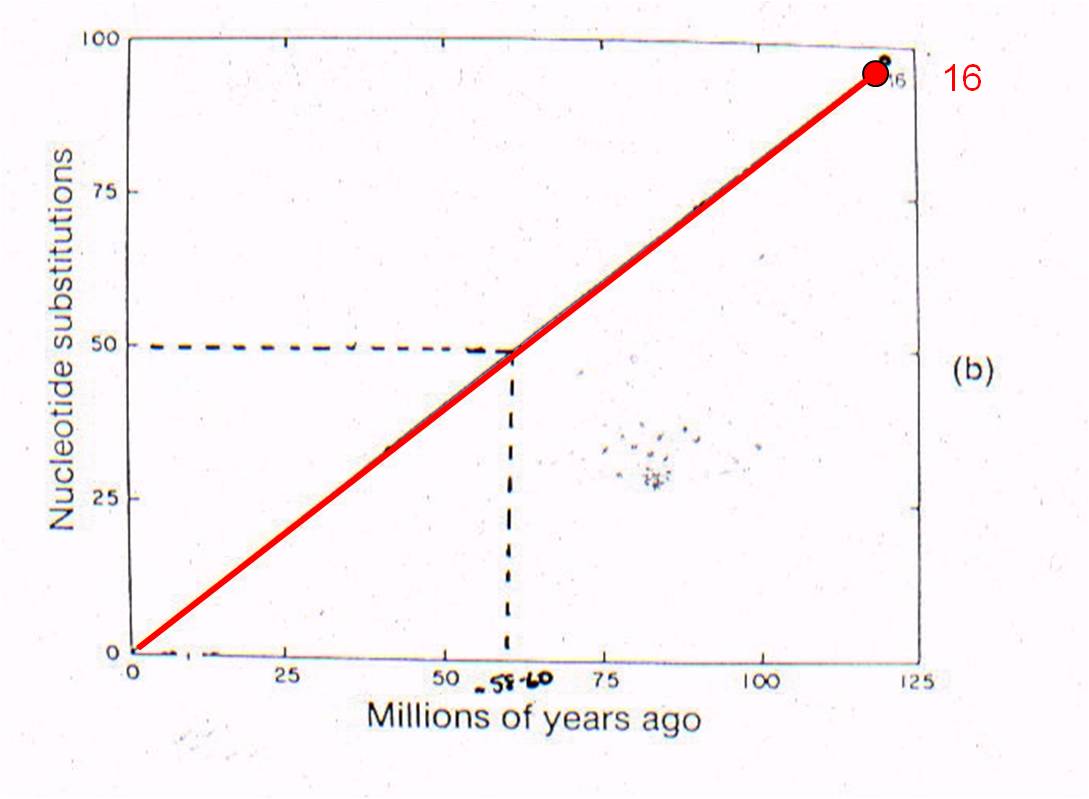
THEN: We can plot 'node 16' based
on the age of the oldest mammal fossil (120 mya) and the genetic difference
measured between marsupials and placentals (98 substitutions in DNA).
AND: if mutation rate
is assumed to be constant, we can draw a straight line from 'node 16' to the
origin. This is the predicted relationship between genetic similarity
and time - predicted by the theory of evolution by common descent and the assumption
of a constant mutation rate. So, evolutionary theory predicts that, if two mammals
differ by 50 substitutions in these seven proteins, then it must have taken
58-60 million years for these differences to accumulate. In other words,
their common ancestor should have lived 58-60 million years ago.
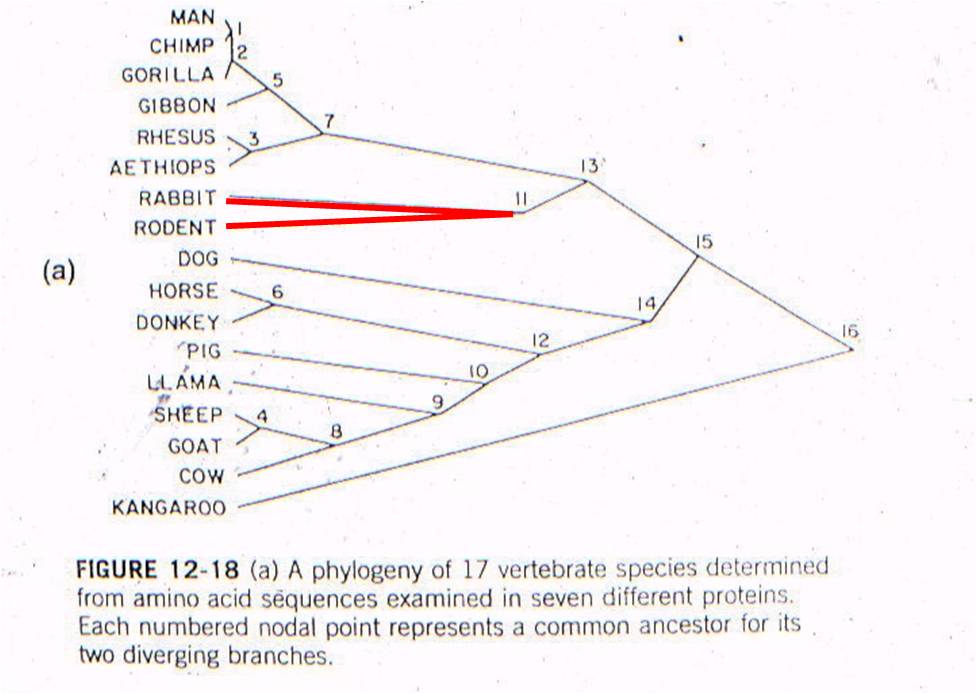
Well, rabbits and rodents differ
by 50 substitutions in the DNA. Our model predicts that the common ancestor
should live 58-60 million years ago. Well, where ARE the presumed ancestors
in the fossil record? They are in strata that date to 58-60 million years
old - just where the genetic analysis of LIVING species predicts they should
be (see node 12, below).
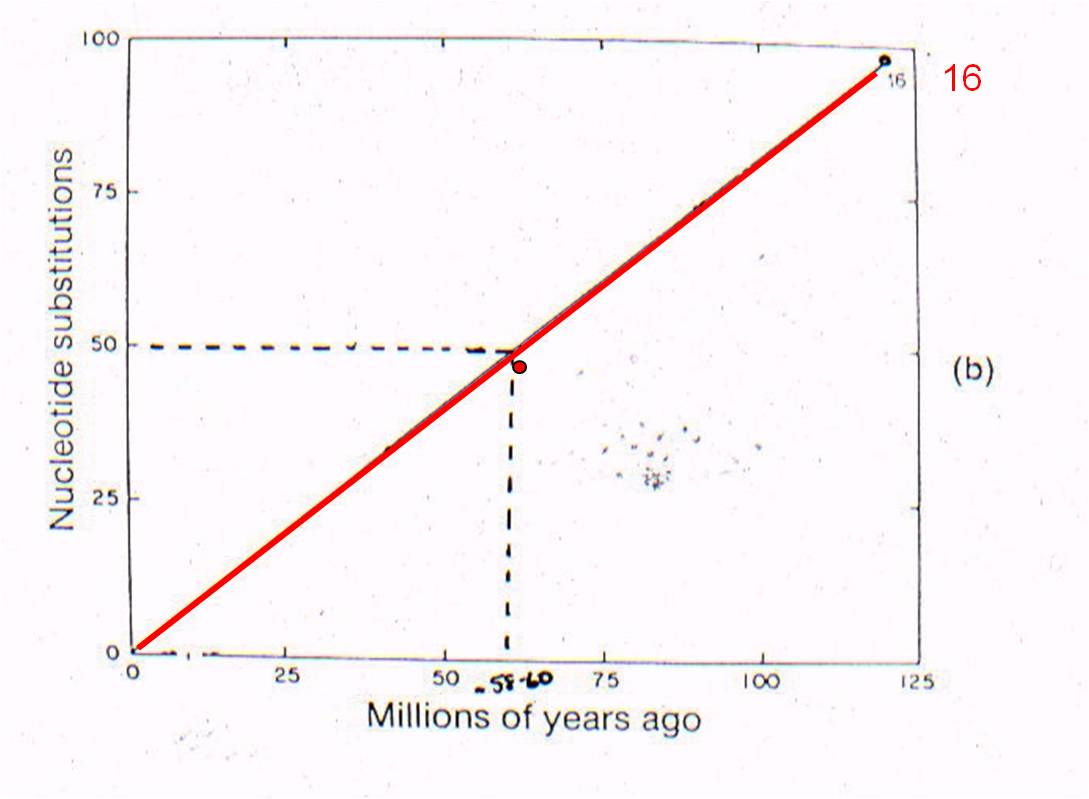
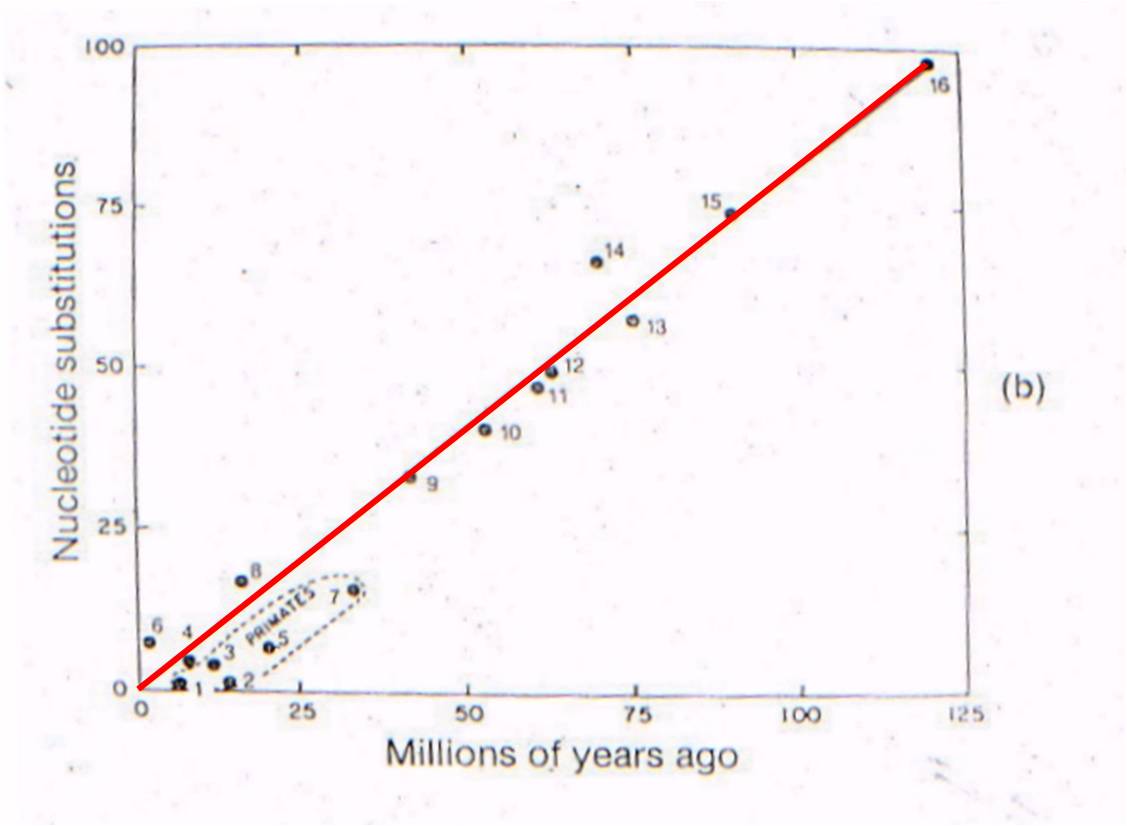 Now
lets consider where all the ancestral fossils are (figure to the right). The
intermediate fossils that link these taxa, and represent these numbered nodes,
are pretty much where our genetic analysis of existing species predicts they
should be - very close to the line. There is some variation - not all
points are exactly on the line - but our assumption of a constant mutation rate
is probably not explicitly correct for all genes, and probably introduces some
slight source of error. None the less, the data is strongly supportive
of our hypothesis - our evolutionary prediction has been confirmed by the data.
Now
lets consider where all the ancestral fossils are (figure to the right). The
intermediate fossils that link these taxa, and represent these numbered nodes,
are pretty much where our genetic analysis of existing species predicts they
should be - very close to the line. There is some variation - not all
points are exactly on the line - but our assumption of a constant mutation rate
is probably not explicitly correct for all genes, and probably introduces some
slight source of error. None the less, the data is strongly supportive
of our hypothesis - our evolutionary prediction has been confirmed by the data.
So, as we have seen before, evolution
not only predicts the existence of common ancestors, but genetic analyses
of living species can predict WHEN, millions to hundreds of millions
of years ago, these different, extinct, ancestral species lived. (Remember
those intermediate fossils? It's not just that they have a combination
of traits, but they existed at the right time. Now we see genetics showing
the same thing, in a PREDICTIVE way, like an good scientific theory should).
The only rational explanation that
explains our ability to do this is evolution from common ancestors. This
wouldn't work if evolution was false, radiaoctive dating was false, or genetic
analyses did not reflect biological relatedness. All these hypotheses
are confirmed by these experiments. Evolution is a predictive, explanatory model
for how the universe works. It has been tested and supported in an extraordinary
variety of ways.
Study Questions:
1. What observations
did Hutton make, and what did he conclude from these observations?
2. What two patterns
occur in the fossil record that impress Darwin regarding the hypothesis of evolution
and common descent?
3. What are homologous
structures? What correlations occurs with the environment?
4. What are analogous
structures? What correlation occurs with the environment?
5. How did
Darwin explain the existence of 'convergent communities"?
6. The Galapagos
are dominated by many unique species of reptiles, birds, and marine mammals. What did this
non-random assemblage suggest to Darwin about their origin, and how was evolution
implied?
7. Why were
the mockingbirds so critical to Darwin's ideas about the production of new species?
8. How did Darwin
use the comparative method and observations of 'artificial selection' to produce
the theory of 'natural selection'?
9. How were Malthus's
observations and conclusion rlevant to the development of Darwin's theory?
10. Outline the
theory of natural selection as an argument, with three premises, 3 conclusions,
and a corollary.
11.
If a rock has a ratio of Ar:K of 7:1, how old is it?
12.
What are the two key characteristics of transitional fossils?
13.
Why is Ichthyostega considered to be an intermediate fossil?
14.
What characteristics make Archaeopteryx an intermediate fossil?
15.
What characteristics do therapsids have that make them intermediate fossils?
16.
What characteristics do Australopithecines have that make them intermediate
fossils?
17.
Explain the logic of using genetic differences and mutational clocks to determine
the time since a common ancestor.
Study
Questions:
11.
What are the five assumptions of the Hardy-Weinberg Equilibrium Model?
12.
Consider the following population:
| |
AA |
Aa |
aa |
| Number
of Individuals |
60 |
20 |
20 |
- -
calculate the genotypic frequencies.
- -
calculate the gene frequencies
- -
calculate the HARDY WEINBERG EQUILIBRIUM frequencies.
13.
If the HWE model does not describe any real population, how can it be useful?