Evolution 1: How Does Life Change?
I. The Growth of Biological Thought
The idea of observing a natural phenomenon,
proposing a testable hypothesis of causality to explain that phenomenon, and
then testing that hypothesis to determine its validity has NOT been a formal
method of inquiry throughout human history. Although processes of "trial
and error" in solving mechanical problems almost necessitate at least an
unconscious 'scientific' process, explanations of how the world IS,
or why it is THIS WAY, were promoted by philosophers long before the
tool of science developed. A very brief overview of the birth of science, as
it relates to the study and explanation of life, is therefore important and
instructive for understanding why Darwin's ideas were both revolutionary and
yet - in some sense - historically anticipated.
A. Western Philosophy and Science
 1.
Plato (427-347 bc):
Plato was trained in the Pythagorean school, and was more truly a pure philosopher
rather than a 'naturalist', per se. As such, he was more impressed by generalizations
rather than the vagarities and variation of individual experience; indeed, those
variations that are so important to a true understanding of biology.
1.
Plato (427-347 bc):
Plato was trained in the Pythagorean school, and was more truly a pure philosopher
rather than a 'naturalist', per se. As such, he was more impressed by generalizations
rather than the vagarities and variation of individual experience; indeed, those
variations that are so important to a true understanding of biology.
UNIVERSAL PHILOSOPHY (four dogmas)
- Essentialism: (essences perfect; material world is an imperfect reflection
of these perfect essences/'ideals' (eidos). Cave analogy; triangle example.
- Universal Harmony: essences form a perfect whole - cant change
- Demi-Urge:
creative force that made the essences
- Soul: non-corporeal "spark of life"
in living systems; maximized in humans.
- became the bedrock of western
civilization for 2000 years! Ernst Mayr, one of the most important biologists
of the 20th century, states: "It took more than 2000 years for biology, under
the influence of Darwin, to escape the paralyzing grip of essentialism...the
rise of modern biology is, in part, the emancipation from Platonic thinking".
2. Thomas
Aquinas (1225-1274) - Aquinas presented the most formal logical argument
for the existence of God, largely using the teleological argument of design.
Events or objects that move towards a goal (have a purpose) a primary cause;
Aristotle's "unmoved mover" is Thomas's Christian God. Thomas professed
a "natural theology", which suggested that one could come to know
more of God by studying "His works" (nature).
 3.
Astronomers: Copernicus, Kepler, Galileo, and Newton: 1543: The publication of Nicolaus
Copernicus's De
revolutionibus orbium coelestium (On the Revolutions of the Heavenly
Spheres) described the heliocentric model of the solar system, opposing the
terracentric view supported by both the authority of the ancients (Ptolemy
and Aristotle) and the Bible. Interestingly, Copernicus still depended on philosophical
preferences over observation - he imagined that the planets travelled in circles,
not ellipses, because the circle was a more perfect form. In short, as Francis
Bacon (1561-1626) concluded, knowledge is incomplete; it is not all found
in the Bible or the ancient texts, and new knowledge is discovereable by the
process of empirical hypothesis testing.
3.
Astronomers: Copernicus, Kepler, Galileo, and Newton: 1543: The publication of Nicolaus
Copernicus's De
revolutionibus orbium coelestium (On the Revolutions of the Heavenly
Spheres) described the heliocentric model of the solar system, opposing the
terracentric view supported by both the authority of the ancients (Ptolemy
and Aristotle) and the Bible. Interestingly, Copernicus still depended on philosophical
preferences over observation - he imagined that the planets travelled in circles,
not ellipses, because the circle was a more perfect form. In short, as Francis
Bacon (1561-1626) concluded, knowledge is incomplete; it is not all found
in the Bible or the ancient texts, and new knowledge is discovereable by the
process of empirical hypothesis testing.
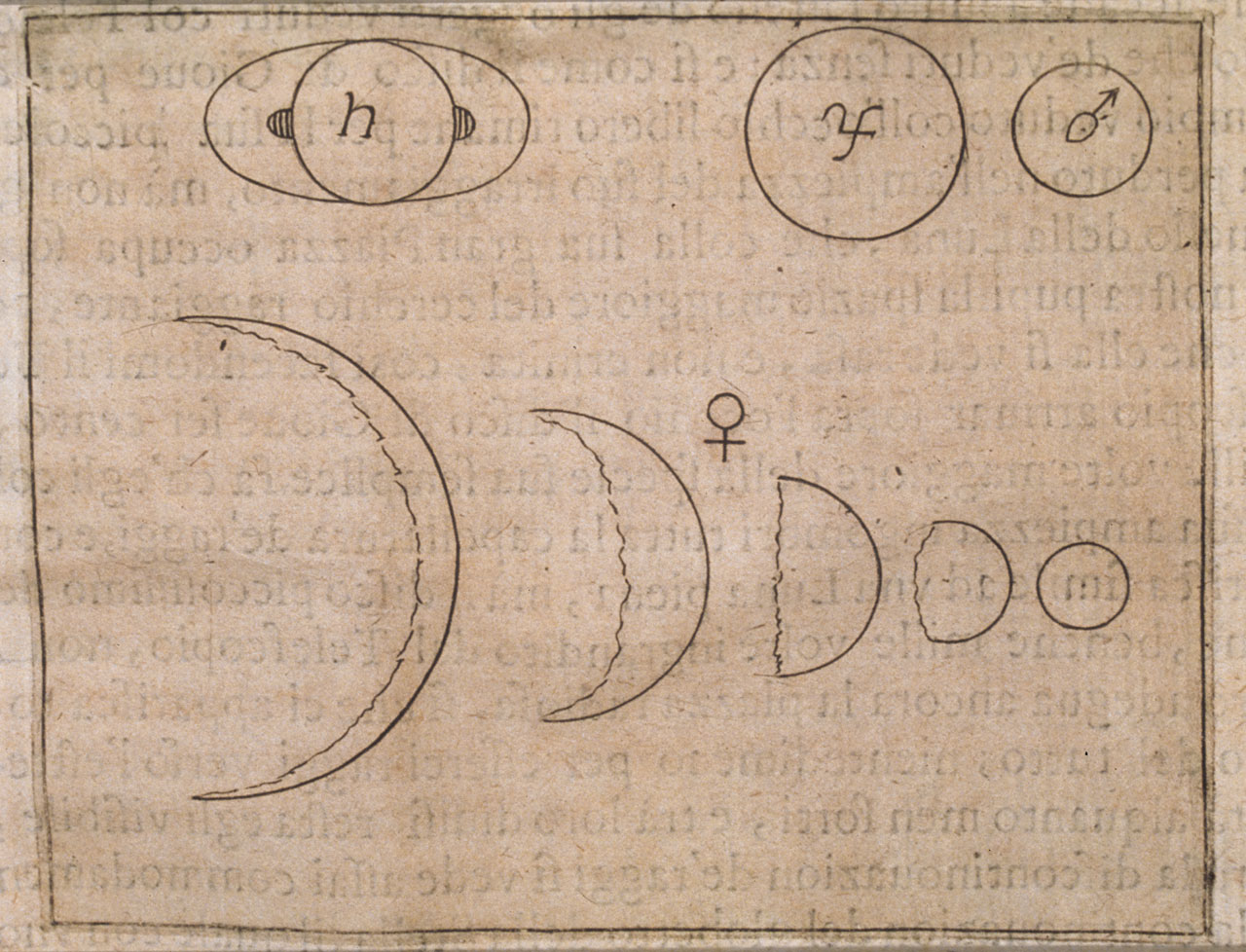 Kepler (1571-1630) and
Galileo (1564-1642)
were the first great natural philosophers in the west to emphasize and use a
mathematical, experimental approach to answer questions about the physical world.
Galileo's observations of the moons orbiting Jupiter, and the full phases
of Venus (impossible to explain with a Ptolemaic model of the solar system),
provide empirical support for the copernican
model. Galileo, however, was still wedded to the philosophical imperative
that the orbits were perfect circles. Kepler attacked this view with voluminous
data collected by he and his mentor, Tycho
Brahe. By using elliptical orbits, Kepler was able to fashion the most precise
predictive models of planetary orbits available. Galileo was a devout Catholic,
but he famously said that the Bible tells a person "how to go to heaven,
not how the heavens go". In this, and in his goad to "Measure what
is measurable, and make measurable what is not so", he personifies the
quantitative scientist. In his Dialogue
Concerning the Two Chief World Systems (1632), he publicized the debate
over these worldviews. The Roman-Catholic church brought him to trial
for heresy, and he was ultimately forced to recant
his support for the Copernican model before the Commissary-General of the Inquisition
in Rome during 1633. He was ultimately placed under house arrest (confined to
his home
in Arcetri) for the remainder of his life.
Kepler (1571-1630) and
Galileo (1564-1642)
were the first great natural philosophers in the west to emphasize and use a
mathematical, experimental approach to answer questions about the physical world.
Galileo's observations of the moons orbiting Jupiter, and the full phases
of Venus (impossible to explain with a Ptolemaic model of the solar system),
provide empirical support for the copernican
model. Galileo, however, was still wedded to the philosophical imperative
that the orbits were perfect circles. Kepler attacked this view with voluminous
data collected by he and his mentor, Tycho
Brahe. By using elliptical orbits, Kepler was able to fashion the most precise
predictive models of planetary orbits available. Galileo was a devout Catholic,
but he famously said that the Bible tells a person "how to go to heaven,
not how the heavens go". In this, and in his goad to "Measure what
is measurable, and make measurable what is not so", he personifies the
quantitative scientist. In his Dialogue
Concerning the Two Chief World Systems (1632), he publicized the debate
over these worldviews. The Roman-Catholic church brought him to trial
for heresy, and he was ultimately forced to recant
his support for the Copernican model before the Commissary-General of the Inquisition
in Rome during 1633. He was ultimately placed under house arrest (confined to
his home
in Arcetri) for the remainder of his life.
 Newton (1642-1727):
In Newton's Philosophiae
Naturalis Principia Mathematica (1687) we see the fulfillment of the
scientific method - the formation of testable general theory. Newton constructed
a general theoretical model of gravity and motion that became classical mechanics.
This theory explained the motion of earthly objects (apples and projectiles
falling) and the elliptical path of heavenly bodies. We see the culmination
of Aristotle's imperative for both inductive and deductive reasoning - from
specific observations one constructs a general hypothesis (inductive reasoning).
Now, you use deduction to create a prediction that follows from that hypothesis
(IF... THEN...). And of course, you subject your prediction to an experimental
test in which falsification is possible. Although other natural philosophers
(the term "scientist" was not coined until the 1830's) had been held
in high regard by some nations, kings, or patrons, Newton was knighted
- signifying the complete cultural acceptance of this new way of examining the
physical world.
Newton (1642-1727):
In Newton's Philosophiae
Naturalis Principia Mathematica (1687) we see the fulfillment of the
scientific method - the formation of testable general theory. Newton constructed
a general theoretical model of gravity and motion that became classical mechanics.
This theory explained the motion of earthly objects (apples and projectiles
falling) and the elliptical path of heavenly bodies. We see the culmination
of Aristotle's imperative for both inductive and deductive reasoning - from
specific observations one constructs a general hypothesis (inductive reasoning).
Now, you use deduction to create a prediction that follows from that hypothesis
(IF... THEN...). And of course, you subject your prediction to an experimental
test in which falsification is possible. Although other natural philosophers
(the term "scientist" was not coined until the 1830's) had been held
in high regard by some nations, kings, or patrons, Newton was knighted
- signifying the complete cultural acceptance of this new way of examining the
physical world.
This "scientific revolution" had a curious effect
on the study of life. Science emancipated physics, astronomy, and chemistry
from theology by describing constant, natural, predictive laws and by describing
the unchanging nature of elements (alchemy disproven). This confirmed the Platonic
views of an unchanging universe created in perfection and left to run like a
'clockwork'. But what about our little corner of the universe? Was the Earth
also static from 'the beginning", and just how long ago was that, anyway?
Anglican Bishop James Ussher (1581-1656) applied logical rigor to the History
of the Earth as revealed in the Bible and counted back the generations, determining
that creation began at noon, October 23, 4004 b.c. (For a great book about calendars,
read Stephen J. Gould's Questioning the Millenium). Thomas Burnet (~1635-1715)
wrote The Sacred Theory of the Earth (1680), an account of earth's
history as a literal account of Genesis 1. So, both concluded that the Earth
was young, and most natural philosphers also concluded that the species were
fixed since their creation.
4. Natural Theologians
- Following on the thinking of Aquinas, there was a resurgence of Natural Theology
to explicitly consider the theological import and relvance of the new observations
made by science. The fundamental assumption was that God made things for a purpose,
and that we might understand God's purpose if we more fully describe the creation
and its operation. The most explicit reconstruction of these ideas was by theologian
William Paley's
Natural Theology: or, Evidences of the Existence and Attributes of the Deity,
Collected from the Appearances of Nature (1802). Here he provides his teleological
argument for the existence of God, using the allegory of the "watchmaker".
 a.
Carl Linne
(1707-1778) - "Linneaus"
(he latinized his own name) was the "great cataloguer", and he published
the first edition of Systema Naturae in 1735. It wasn't until 1753,
however, in Species Plantarum (Plant Species), that he formalized his prcedure
for using two names to identify a species - the latin binomen (like Homo
sapiens). The first name is the GENUS, and the second is called the 'specific
epithet' that describes this species and distinguishes it from other similar
species placed in the same genus. Linnaeus also grouped these genera (plural
of 'genus') into orders, classes, and kingdoms based on additional morphological
similarities. In plants, he relied on similarities in the reproductive structures,
as many naturalists accepted that species are kinds that reproduce only with
themselves. In the 10th edition of Systema Naturae (1758), he applied
this system to all animals, too. The oldest scientific species names used today
date from these two works. By the way, the word 'species' is both the singular
and plural. There is no 'specie'. FYI. : )
a.
Carl Linne
(1707-1778) - "Linneaus"
(he latinized his own name) was the "great cataloguer", and he published
the first edition of Systema Naturae in 1735. It wasn't until 1753,
however, in Species Plantarum (Plant Species), that he formalized his prcedure
for using two names to identify a species - the latin binomen (like Homo
sapiens). The first name is the GENUS, and the second is called the 'specific
epithet' that describes this species and distinguishes it from other similar
species placed in the same genus. Linnaeus also grouped these genera (plural
of 'genus') into orders, classes, and kingdoms based on additional morphological
similarities. In plants, he relied on similarities in the reproductive structures,
as many naturalists accepted that species are kinds that reproduce only with
themselves. In the 10th edition of Systema Naturae (1758), he applied
this system to all animals, too. The oldest scientific species names used today
date from these two works. By the way, the word 'species' is both the singular
and plural. There is no 'specie'. FYI. : )
 b.
Georges Louis Leclerc,
Comte de Buffon (1707-1788)
Buffon pulbished the first volume of his encyclopedic Histoire Naturelle
in 1749. Ernst Mayr considered Buffon to be the foremost biologist of the 18th
century, and Mayr wrote "it makes no difference which of the authors of the
second half of the 18th century one reads - their discussions are, in the last
analysis, merely commentaries on Buffons work. Except for Darwin and Aristotle,
there has been no other student of organisms who has had as far-reaching an
influence." He opposed the notion oif classification; if the species were separately
created, then of what use was any classification system? He was aware of possibility
of evolution but rejects it:
b.
Georges Louis Leclerc,
Comte de Buffon (1707-1788)
Buffon pulbished the first volume of his encyclopedic Histoire Naturelle
in 1749. Ernst Mayr considered Buffon to be the foremost biologist of the 18th
century, and Mayr wrote "it makes no difference which of the authors of the
second half of the 18th century one reads - their discussions are, in the last
analysis, merely commentaries on Buffons work. Except for Darwin and Aristotle,
there has been no other student of organisms who has had as far-reaching an
influence." He opposed the notion oif classification; if the species were separately
created, then of what use was any classification system? He was aware of possibility
of evolution but rejects it:
"Not only the ass and the horse, but also man, the apes, the quadrupeds,
and all the animals might be regarded as constituting but a single family...
If it were admitted that the ass is of the family of the horse, and different
from the horse only because it has varied from the original form, one could
equally well say that the ape is of the family of man, that he is a degenerate
man, that man and ape have a common origin; that, in fact, all the families,
among plants as well as animals, have come from a single stock, and that all
the animals are descended from a single animal, from which have sprung in the
course of time, as a result of progress or of degeneration, all the other races
of animals. For if it were once shown that we are justified in establishing
these families; if it were granted that among animals and plants there has been
(I do say several species) but even a single one, which has been produced in
the course of direct decent from another species; if, for example, it were true
that the ass is but a degeneration from the horse - then there would no longer
be any limit to the power of nature, and we should not be wrong in supposing
that, with sufficient time, she has been able from a single being to derive
all the other organized beings. But this is by no means a proper representation
of nature. We are assured by the authority of revelation that all animals have
participated equally in the grace of direct Creation and that the first pair
of every species issued forth fully formed from the hands of the Creator."
Histoire Naturelle (1753)
 c.
Jean Baptiste Pierre Antoine de Monet, Chevalier de Lamarck
(1744-1829) - Initially
a botanist and tutor to Buffon's son, he became an expert in invertebrates and
the mollusc fossils of the Paris Basin. through this work, his initial belief
in the fixity of species changed. In his culminating work Philosophie
Zoologique (1809), he suggested that species change over time, climbing
the Scala Naturae from simple forms to complex. The simplest forms were continuously
produced by spontaneous generation, and species did not go extinct - they evolved
into more complex forms over long periods of time. In addition to this vertical
progression, they could also diverge as a consequence of responding to the environment
and passing on the traits they acquired as a result of the action of this environment.
Structures that were used in an environment would be elaborated (use and disuse),
and then these newly elaborated structures were passed on to offspring (inheritance
of acquired traits). Lamarck's evolutionary ideas explained the new observations
of fossil diversity and apparent extinction. In Lamarck's mind, extinctions
were theologically impossible, because he believed in a complete, harmonious
creation by a benevolent creator. Why would a benevolent, purposeful creator
let creations go extinct, and wouldn't the loss of some species render the perfection
of the initial creation imperfect? For Lamarck, species changing into other
species preserved the whole of creation. "May it not be possible that the fossils
in question belong to species still existing, but which have changed since that
time and have been converted into that similar species that we now actually
find?" Lamarck is rightly regarded as the first "biologist" (he was
the first to use the term) to propose a true evolutionary hypothesis, and a
testable, naturalistic mechanism to explain it. Unfortunately, the mechanism
was wrong.
c.
Jean Baptiste Pierre Antoine de Monet, Chevalier de Lamarck
(1744-1829) - Initially
a botanist and tutor to Buffon's son, he became an expert in invertebrates and
the mollusc fossils of the Paris Basin. through this work, his initial belief
in the fixity of species changed. In his culminating work Philosophie
Zoologique (1809), he suggested that species change over time, climbing
the Scala Naturae from simple forms to complex. The simplest forms were continuously
produced by spontaneous generation, and species did not go extinct - they evolved
into more complex forms over long periods of time. In addition to this vertical
progression, they could also diverge as a consequence of responding to the environment
and passing on the traits they acquired as a result of the action of this environment.
Structures that were used in an environment would be elaborated (use and disuse),
and then these newly elaborated structures were passed on to offspring (inheritance
of acquired traits). Lamarck's evolutionary ideas explained the new observations
of fossil diversity and apparent extinction. In Lamarck's mind, extinctions
were theologically impossible, because he believed in a complete, harmonious
creation by a benevolent creator. Why would a benevolent, purposeful creator
let creations go extinct, and wouldn't the loss of some species render the perfection
of the initial creation imperfect? For Lamarck, species changing into other
species preserved the whole of creation. "May it not be possible that the fossils
in question belong to species still existing, but which have changed since that
time and have been converted into that similar species that we now actually
find?" Lamarck is rightly regarded as the first "biologist" (he was
the first to use the term) to propose a true evolutionary hypothesis, and a
testable, naturalistic mechanism to explain it. Unfortunately, the mechanism
was wrong.
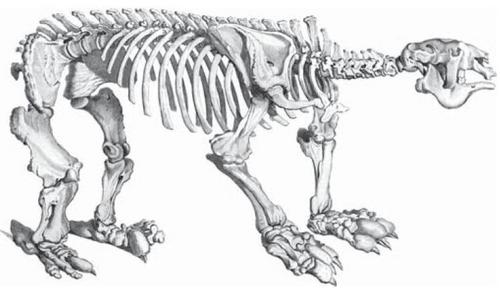 d.
Georges Cuvier
(1769-1832) - Cuvier was also an intellectual giant in France,
and was Lamarck's nemesis. A great anatomist, Cuvier founded the comparative
approach in anatomy and also founded vertebrate paleontology. Unlike Lamarck,
he believed that extinctions occurred; and he supported this contention by showing
that the great mammals of the past (mammoths, giant ground sloths, etc.) had
no modern living forms (Cuvier concluded that elephants and mammoths were different
species, and mammoths were truly extinct.) In contrast to Lamarck's ideas of
gradually changing species responding to their environment, Cuvier promoted
the notion that there were period cataclysms ('revolutions') that killed off
local faunas and required repopulation from elsewhere. This idea became known
as 'catastrophism', and stood in opposition to the 'uniformitarianism' of gradual
change espoused by Lamarck and others. Cuvier also tore down the notion of the
scala naturae, replacing it with four 'embranchments' of life disconnected from
eachother. Finally, Cuvier said that the intermediates predicted by Lamarck's
gradualistic evolutionary model did not exist, and he reminded the scientific
community that spontaneous generation had been refuted (for insects, at least)
since the experiments of Francesco Redi in 1668! He was a true essentialist,
seeing discontinuity through time and through taxa. Cuvier outlived Lamarck
and continued to lambast him and his ideas. Evolutionary ideas fell into disfavor
as a consequence.
d.
Georges Cuvier
(1769-1832) - Cuvier was also an intellectual giant in France,
and was Lamarck's nemesis. A great anatomist, Cuvier founded the comparative
approach in anatomy and also founded vertebrate paleontology. Unlike Lamarck,
he believed that extinctions occurred; and he supported this contention by showing
that the great mammals of the past (mammoths, giant ground sloths, etc.) had
no modern living forms (Cuvier concluded that elephants and mammoths were different
species, and mammoths were truly extinct.) In contrast to Lamarck's ideas of
gradually changing species responding to their environment, Cuvier promoted
the notion that there were period cataclysms ('revolutions') that killed off
local faunas and required repopulation from elsewhere. This idea became known
as 'catastrophism', and stood in opposition to the 'uniformitarianism' of gradual
change espoused by Lamarck and others. Cuvier also tore down the notion of the
scala naturae, replacing it with four 'embranchments' of life disconnected from
eachother. Finally, Cuvier said that the intermediates predicted by Lamarck's
gradualistic evolutionary model did not exist, and he reminded the scientific
community that spontaneous generation had been refuted (for insects, at least)
since the experiments of Francesco Redi in 1668! He was a true essentialist,
seeing discontinuity through time and through taxa. Cuvier outlived Lamarck
and continued to lambast him and his ideas. Evolutionary ideas fell into disfavor
as a consequence.
1. Observations
a. The Earth is old: James
Hutton (1726-1797): Hutton was the first great british geologist.
He compared Hadrian's wall - which looks new but was 1600 years old (122 AD)
- with natural rock outcrops that were strongly weathered. Hutton concluded
that the natural outcrops must be 100's of times older. He also examined an
important formation at Siccar Point, where one series of nearly vertical strata
is overlain by another series of horizontal strata. This is now called an 'unconformity',
and Hutton explained it as follows. Based on Steno's laws of superposition,
the bottom vertical sediments must have been laid down first, and they must
have been laid down horizontally. Ages must have passed between each deposit,
as each turned to rock. Then, uplifts must have occurred to bend them into a
vertical aspect. Long periods of erosion must take place to wear that uplift
flat, followed by the long intervals of time needed to deposit the second horizontal
series. Also, if erosion and deposition acted slowly (as current observations
show), then it must have taken a really long time to erode mountains or build
up marine deposits (White Cliffs of Dover). He concluded that this slow, 'uniformitarian'
cycle of deposition, uplift, erosion, and deposition meant that the Earth was
unfathomably old. Indeed, the cycle may mean that it's age might not be discoverable.
In short, Hutton concludes, the Earth has "no vestige of a beginning, no prospect
of an end."
b. Paleontology: Fossils show Patterns
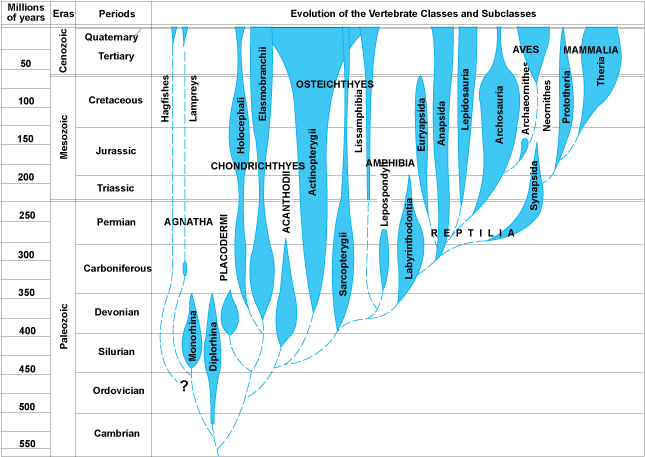 Paleontology
provided a variety of interesting patterns. First, there were extinct forms
that were different from the species alive today. Although some earlier natural
philosophers suggested that the creatures might still exist in some unexplored
corner of the globe, that was a less satisfying hypothesis in the mid-1800's...
most areas of the globe had been visited by Europeans. Also, the idea of extinction
was repugnant to some people on theological grounds. If God had created a perfect
world, then extinction renders that creation imperfect. Also, if species could
go extinct since the creation, could species also come into existence since
the creation? Just how dynamic was this system?
Paleontology
provided a variety of interesting patterns. First, there were extinct forms
that were different from the species alive today. Although some earlier natural
philosophers suggested that the creatures might still exist in some unexplored
corner of the globe, that was a less satisfying hypothesis in the mid-1800's...
most areas of the globe had been visited by Europeans. Also, the idea of extinction
was repugnant to some people on theological grounds. If God had created a perfect
world, then extinction renders that creation imperfect. Also, if species could
go extinct since the creation, could species also come into existence since
the creation? Just how dynamic was this system?
Darwin was impressed by two major
patterns in the fossil record.
1. The major groups of animals accumulate
in an orderly manner'. Everything is not represented at the beginning. In vertebrates,
for instance, the fishes appear first, and exist throughout the rest of the
record. Amphibians appear next, followed by reptiles, mammals, and birds. So
it is not everything at the beginning, and it is not a replacement. Where did
mammals come from? Spontaneous generation had been refuted, so Darwin knew that
mammals had to come from other pre-existing animals. But the only completely
terrestrial vertebrates before mammals were reptiles.
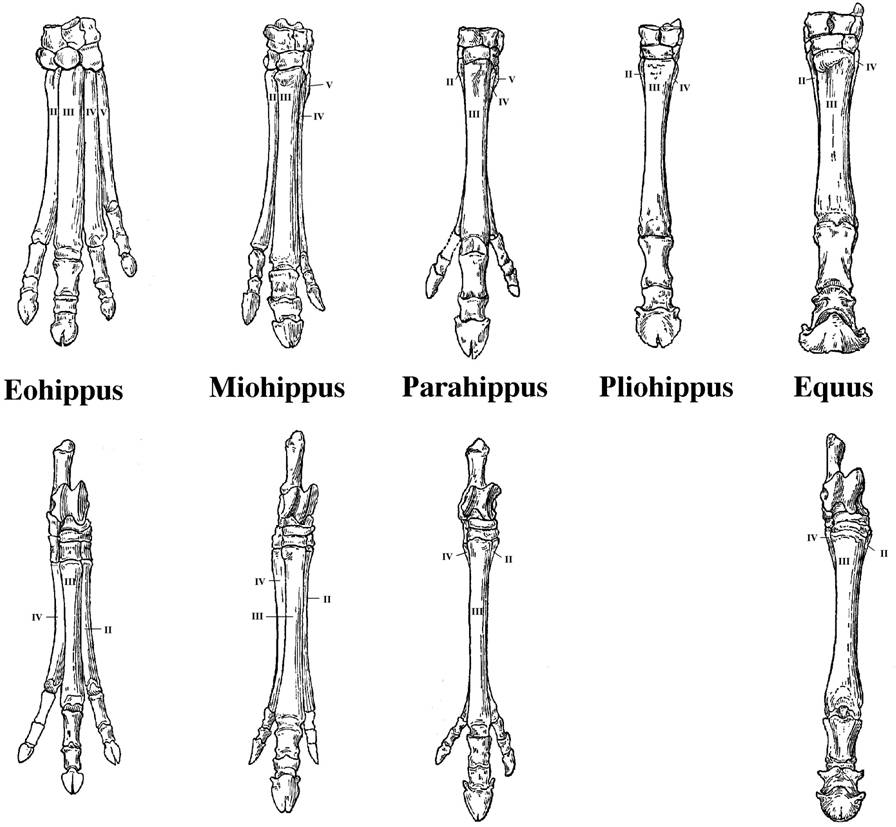
2. A second major pattern occurred
within some lineages of similar organisms. Within some lineages, we seen orderly
change in the size or characteristics of species in a geological sequence. For
instance, consider the morphological patterns in a particular taxon (horses).
Fossils in a stratigraphic sequence are similar, but often have traits that
form a continuum...like the progressive loss of digits on the horse limb. And,
with each innovation, there are often radiations - a "spurt" in the
number of species that show this new trait. And finally, these species in recent
strat are more similar to living ('extant') species than the species found in
deeper, older strata. So, many of these transitional sequences terminate in
living representatives.

 c.
Comparative Anatomy shows patterns realted to the environment:
c.
Comparative Anatomy shows patterns realted to the environment:
Homologous Structures
Although having a different outward "look" and although used
for different purposes, they have an underlying similarity in structure - forelimbs
of vertebrates all have one long upper arm bone, two lower arm bones, a bunch
of wrist bones, and five digits. Darwin saw the similarity in structure as important.
An engineer builds different things for different purposes - cars, boats, and
airplanes are structurally DIFFERENT. Here, however, it seemed as if one basic
structure was modified for different uses. Darwin knew why siblings in a family
were similar - they had the same parents (ancestors). He reasoned that these
structural similarities in different species might be due to the same principle
- common ancestry. Also, he observed a correlation: Different uses correlated
with different environments. Could this correlation be causal?
Analogous
Structures
Organisms in the same environment often have a similar outward
structure or body plan. For example, flying animals all have an aerodynamic
wing that is wider at the front than at the rear. However, the wings of differnt
animals are differnt in underlying structure. Bats have fingers that support
the membraneous wing, whereas birds lack fingers and the body of the wing consists
of feathers. Insect wings don't involve the limbs at all (even though they have
6!). Again, Darwin observed this correlation with the environment: similar use
(and outward structure) in similar environments. Could this correlation be causal?
Vestigial Organs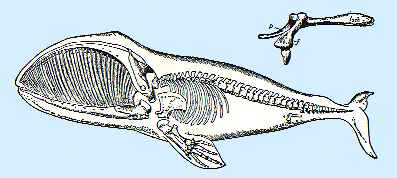
These are organs that have no function in one organism (where
they are 'vestigial') but they do function in other organisms. So, some whales
have hip bones, but no legs. Why do they have these bones? Darwin was struck
by the IMPERFECTIONS in nature, as much as the adaptations. Why do men have
nipples? Why do we have muscles that wiggle our ears? Why do we have strong
muscles in the front of our stomach, which are not "load-bearing", and weak
muscles at the base of our abdomen (which rupture in a hernia)? This is a reasonable
relationship in a quadraped, but not in a biped. Why do we have tail bones,
but no external tail? Again, these are NOT well-designed features. In fact,
attributing these imperfect designs to a perfect creator could be interpreted
as heretical. However, when we see them working in OTHER species, it suggests
that maybe we inherited them from common ancestors where they DID serve a function.
As a scientist, Darwin was trying to explain ALL the data (adaptations and imperfections),
he was not simply bringing forward only the data that supported a preferred
position (design).
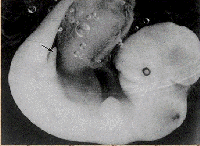
d. Species distributions show patterns realtd to the environment:
Under similar environmental conditions, we find different
species filling similar ecological niches. Outward 'form' correlates with
ecological niche (role) across entire communities. So, in Australia, marsupials
fill the role of dog-like predator, cat-like predator, burrowing animal, ant-eater,
etc. These same roles are filled by outwardly similar placental mammals in South
America. However, the similarity between a wolf (placental) and a Thylacine
(marsupial - the 'tasmanian wolf') are strictly ANALOGIES. Their underlying
structure shows them to be quite different - a wolf is more similar to a ground
hog (both placentals) in underlying structure than to a thylacine. 
Islands often
have fewer species than a mainland - even a patch of mainland the same size.
As such, the patterns and interactions are often simpler to describe and understand.
For both Darwin and Alfred Russel Wallace (the other independent author of the
theory of evolution by natural selection), the study of islands was critical
in to the development of their ideas.
1. Distance correlates with the
uniqueness of the inhabitants: the animals on the Fauklands are the same
species as on the mainland, but the Galapagos fauna is composed of unique species,
found nowhere else:
"The natural history of these islands is eminently curious, and well deserves
attention. Most of the organic productions are aboriginal creations, found nowhere
else; there is even a difference between the inhabitants of the different islands;
yet all show a marked relationship with those of America, though separated from
that continent by an open space of ocean, between 500 and 600 miles in width.
The archipelago is a little world within itself, or rather a satellite attached
to America, whence it has derived a few stray colonists, and has received the
general character of its indigenous productions. Considering the small size
of the islands, we feel the more astonished at the number of their aboriginal
beings, and at their confined range. Seeing every height crowned with its crater,
and the boundaries of most of the lava- streams still distinct, we are led to
believe that within a period geologically recent the unbroken ocean was here
spread out. Hence, both in space and time, we seem to be brought somewhat near
to that great fact -- that mystery of mysteries -- the first appearance of new
beings on this earth." The Voyage of the Beagle - Darwin (1839).
 2. The Galapagos fauna:
2. The Galapagos fauna:
- It was related to american fauna,
yet different: the types of animals are new world animals.... there are iguanas
like the green iguana of Central and South America, but the iguanas are different
species. So, darwin describe it as " a world within itself, or rather,
a satellite of the Americas" .... it was different, but more like the American
fauna than any other...(no chameleons, for instance, which are old world lizards...)
- It was dominated by dispersive forms. This is critical. The communities are
dominated by reptiles, birds, and marine mammals. All of these organisms could
MIGRATE to the islands from the mainland. (Terrestrial mammals don't migrate
as well as terrestrial reptiles over open ocean. Throw a reptile in cold salty
water, and: 1) its metabolism slows down (its cold), so 2) its demand for food
and water decline; and 3) its scales protect it against water loss... which
is why reptiles do well in the desert, too. Throw a mammal in cold salt water,
and it's going to have a VERY tought time: 1) the temperature gradient between
its warm body and the cold ocean is very large - in order to maintain its high
body temperature against this gradient, it's metabolism has to INCREASE (to
produce more heat to compensate for the heat lost to the environment). This
increased metabolic demand will INCREASE the need for food and water... that's
probably in pretty short supply in the open ocean; and 2) water is lost quickly
from the skin to the salty ocean once the fur is wet... so, mammals are more
likely to starve or die of exposure than reptiles.
-
So, the islands are dominated by dispersive forms, and this suggests they came
from America. But if they came from America, WHY ARE THEY DIFFERENT SPECIES
THAN THOSE IN AMERICA? They must have changed since their arrival.
- There are even differences between
species on different islands. On the 14 species of finches - "Seeing this gradation
and diversity of structure in one small, intimately related group of birds,
one might really fancy that from an original paucity of birds in this archipelago,
one species had been taken and modified for different ends." The Voyage
of the Beagle - Darwin (1839) VIDEO
2. Argument
For Evolution as a Historical Fact:
Premise 1: Species
that are alive today are different from those that have lived previously.
Premise 2: Spontaneous Generation is refuted, so organisms
only come from other organisms.
Conclusion 1: Thus, the organisms alive today must have come
from those pre-existing, yet different, species.
Conclusion 2: There must have been change through time (evolution).
Conclusion 3: The fossil record, vestigial organs, and homologies
are all suggestive of descent from common ancestors.
Below, the figure from The Origin
of Species that shows Darwin's idea of descent from common ancestors.
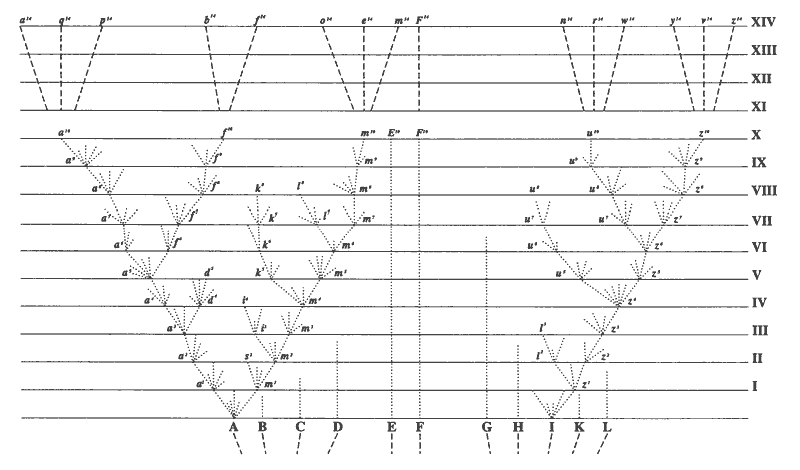
So, if species do change over time
(evolve), the next question is "How?" How does this change occur?
3. How does change occur? Natural Selection
(Know this. Understand it. You WILL be asked to outline NS in this
very form.)
P1: Populations over-reproduce (Malthus)
P2: resources are finite (Malthus)
C1: Eventually, a population will grow until it becomes limited by its resources.
At that time, their will be a "struggle for existence" and most offspring
produced will die. (Malthus)
P3: Individuals in a population vary, and some of this variation is heritable (Darwin
- observations and animal/plant breeding)
C2: Variations will not have the same probability of survival and reproduction in
a particular environment; those well-suited to the environment will be more
likely to survive and reproduce than others, passing on the genes for these
adapted traits. There will be "Differential Reproductive Success" (Observations,
breeding).
C3: Over time, adaptive traits will accumulate and the characteristics in a population
will change. This is lineage evolution. (Like change in horse toes in a sequence
of fossil species, or like the change in the chihuahua lineage from the ancestral
wolves).
Corollary: Two sub-populations, separated in different environments, would be selected
for different traits and may subsequently lose the capacity to interbreed. At
this point, they are different biological species. This is Speciation and Radiational
Evolution. (like the production of different Finches, mockingbirds, etc. on
different islands in the galapagos, and like the radiation of St. Bernards AND
chihuahua's, which diverged from one another over time).
Darwin here provides a natural explanation
for why purposeful structures and behaviors occurs in nature. Through some process
unknown to him, variation arises in natural populations. These varieties differ
in terms of functional efficiency in a common environment; so some improving
an organisms probability of surviving and mating than others. Organisms with
these beneficial traits will leave more offspring, and the frequencies of these
beneficial characteristics will increase through time - much as humans select
for smaller and smaller dogs. He ends The Origin of Species (1859)
like this:
 "It
is interesting to contemplate an entangled bank, clothed with many plants of
many kinds, with birds singing on the bushes, with various insects flitting
about, and with worms crawling through the damp earth, and to reflect that these
elaborately constructed forms, so different from each other, and dependent on
each other in so complex a manner, have all been produced by laws acting around
us. These laws, taken in the largest sense, being Growth with Reproduction;
Inheritance which is almost implied by reproduction; Variability from the indirect
and direct action of the external conditions of life, and from use and disuse;
a Ratio of Increase so high as to lead to a Struggle for Life, and as a consequence
to Natural Selection, entailing Divergence of Character and the Extinction of
less-improved forms. Thus, from the war of nature, from famine and death, the
most exalted object which we are capable of conceiving, namely, the production
of the higher animals, directly follows. There is grandeur in this view of life,
with its several powers, having been originally breathed into a few forms or
into one; and that, whilst this planet has gone cycling on according to the
fixed law of gravity, from so simple a beginning endless forms most beautiful
and most wonderful have been, and are being, evolved". - The Origin of Species (Darwin 1859).
"It
is interesting to contemplate an entangled bank, clothed with many plants of
many kinds, with birds singing on the bushes, with various insects flitting
about, and with worms crawling through the damp earth, and to reflect that these
elaborately constructed forms, so different from each other, and dependent on
each other in so complex a manner, have all been produced by laws acting around
us. These laws, taken in the largest sense, being Growth with Reproduction;
Inheritance which is almost implied by reproduction; Variability from the indirect
and direct action of the external conditions of life, and from use and disuse;
a Ratio of Increase so high as to lead to a Struggle for Life, and as a consequence
to Natural Selection, entailing Divergence of Character and the Extinction of
less-improved forms. Thus, from the war of nature, from famine and death, the
most exalted object which we are capable of conceiving, namely, the production
of the higher animals, directly follows. There is grandeur in this view of life,
with its several powers, having been originally breathed into a few forms or
into one; and that, whilst this planet has gone cycling on according to the
fixed law of gravity, from so simple a beginning endless forms most beautiful
and most wonderful have been, and are being, evolved". - The Origin of Species (Darwin 1859).
II. Modern Evolution: Population Genetics
A. Overview
As a consequence of our modern understanding
of heredity and genetics, we have learned quite a bit about variation AND evolution,
and our model, a this point in the class is:
Sources of Variation Agents of Change
MUTATION:
-New Genes:
Natural Selection
point mutation
Mutation (polyploidy can make new species)
RECOMBINATION:
- New Genes:
exon shuffling
-New Genotypes:
-crossing over
- independent assortment
 In
the early 20th century, at the same time that T. H. Morgan was studying mutations
and creating linkage maps, other biologists were considering the evolutionary
implications of this new knowledge regarding genetic variation. They appreciated
that individuals do not evolve - evolution is a process that occurs at the population
level. For example, as a consequence of differential reproductive success among
individuals in a population, the range of phenotypes and their relative frequencies
in the population will change over time. Individuals are born, life, reproduce
(maybe) and die. As a result of passing on their genes at different frequencies,
the genetic structure of the population changes over time (evolution). Two biologists,
G. Hardy and W. Weinberg, constructed a model to explain how the genetic structure
of a population might change over time.
In
the early 20th century, at the same time that T. H. Morgan was studying mutations
and creating linkage maps, other biologists were considering the evolutionary
implications of this new knowledge regarding genetic variation. They appreciated
that individuals do not evolve - evolution is a process that occurs at the population
level. For example, as a consequence of differential reproductive success among
individuals in a population, the range of phenotypes and their relative frequencies
in the population will change over time. Individuals are born, life, reproduce
(maybe) and die. As a result of passing on their genes at different frequencies,
the genetic structure of the population changes over time (evolution). Two biologists,
G. Hardy and W. Weinberg, constructed a model to explain how the genetic structure
of a population might change over time.
Their model begins by constructing
an 'equilibrium' model - a model of what the gentic structure would look like,
and how it would behave, if there was NO CHANGE over time. (We can liken this
to a "stitstical null hypothesis of no effect"). Then, an actual population
is compared to this model, so see whether the population is evolving or not.
B. The Genetic Structure of a Population
Our first step is to describe the
genetic structure of a population; we need to do this before we can model what
it would do over time. The genetic structure of a population is defined by the
gene array and the genotypic array. To understand what these are, some definitions
are necessary:
1. Definitions:
- Evolution: a change in the genetic structure of a population
- Population: a group of interbreeding organisms that share
a common gene pool; spatiotemporally and genetically defined
- Gene Pool: sum total of alleles held by individuals in a population
- Genetic structure: Gene array and Genotypic array
- Gene/Allele Frequency: % of alleles at a locus of a particular
type
- Gene Array: % of all alleles at a locus: must sum to 1.
- Genotypic Frequency: % of individuals with a particular genotype
- Genotypic Array: % of all genotypes for loci considered;
must = 1.
2. Basic
Computations - Determining
the Genotypic and Gene Arrays:
 The
easiest way to understand what these definitions represent is to work a problem
showing how they are computed.
The
easiest way to understand what these definitions represent is to work a problem
showing how they are computed.
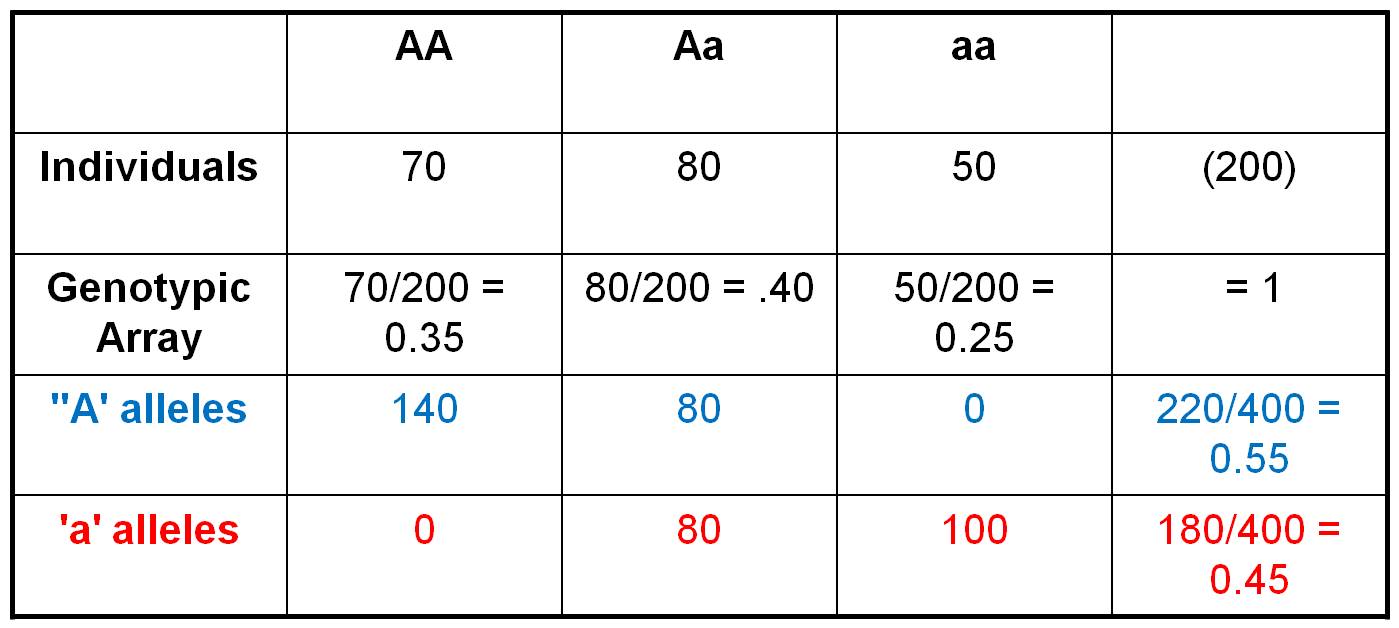
Consider the population shown to
the right, in which there are 70 AA individuals, 80 heterozygotes, and 50 aa
individuals. We can easily calculate the Genotypic Frequencies by dividing each of these values by the total number of individuals in the population.
So, the Genotypic Frequency of AA = 70/200 = 0.35. If we account
for all individuals in the population (and haven't made any careless math erros),
then the three genotypic frequencies should sum to 1.0. The Genotypic
Array would list all three genotypic frequencies: f(AA) = 0.35,
f(Aa) = 0.40, f(aa) = 0.25. A Gene Frequency is the
% of all genes in a population of a given type. This can be calculated two ways.
First, let's do it the most obvious and direct way, by counting the alleles
carried by each individual. So, there are 70 AA individuals. Each carries 2
'A' alleles, so collectively they are 'carrying' 140 'A' alleles. The 80 heterozygotes
are each carrying 1 'A' allele. And of course, the 'aa' individuals aren't carrying
any 'A' alleles. So, in total, there are 220 'A' alleles in the population.
With 200 diploid individuals, there are a total of 400 alleles at this locus.
So, the gene frequency of the 'A' gene = f(A) = 220/400 = 0.55.
We can calculate the frequency of the 'a' alleles the same way. The 50 'aa'
individuals are carrying 2 'a' alleles each, for a total of 100 'a' alleles.
The 80 heterozygotes are each carrying an 'a' allele, and the 140 AA homozygotes
aren't carrying any 'a' alleles. So, in total, there are 180 'a' alleles out
of a total of 400, for a gene frequency f(a) = 180/400 = 0.45. The gene
array presents all the gene frequencies, as: f(A) = 0.55, f(a)
= 0.45.
There is a faster way to calculate
the gen frequencies in a population than adding up the genes contributed by
each genotype. Rather, you can use these handy formulae:
f(A) = f(AA) + f(Aa)/2
f(a) = f(aa) + f(Aa)/2
So, to calculate the frequency of
a gene in a population, you add the frrequency of homozygotes for that allele
with 1/2 the frequency of heterozygotes. In our example, this would be:
f(A) = 0.35 + 0.4/2 = 0.35 + 0.2
= 0.55
f(a) = 0.25 + 0.4/2 = 0.25 + 0.2
= 0.45
Wow... that's alot faster.
C. The Hardy-Weinberg
Equilibrium Model
1. Goal:
The goal of the "Hardy-Weinberg
Equilibrium Model" (HWE) is to describe what the genetic structure of the
population would be if NO evolutionary change occurs. Working independently, Hardy and Weinberg realized
that the gene frequencies in a population will NOT change - will remain in EQUILIBRIUM
- if the following conditions are met:
- there is random mating
- no selection
- no mutation
- no migration
- and the population is infinitely
large.
And, they realized that a population
will reach an equilibrium in GENOTYPIC frequencies, too, after one generation
of meeting these expectations (and for as long as it does so), a population
will NOT EVOLVE. Let's see how they came by these conditions.
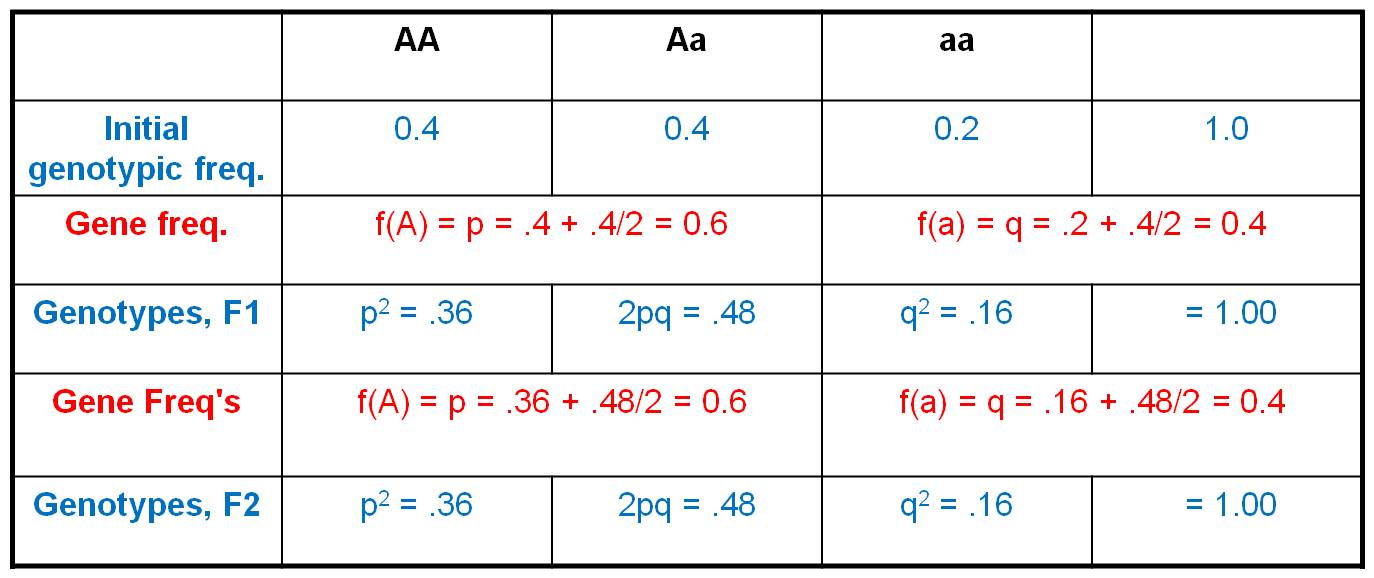 2.
Example:
2.
Example:
Consider the initial population,
with a genotypic array as shown. The gene frequencies are:
A = 0.4 + (0.4/2) = 0.6
a = 0.2 + (0.4/2) = 0.4
Now, consider this gene pool in which
60% of the alleles are 'A' and 40% of the alleles are 'a' (as defined by the
gene frequencies).
So, now we employ the HWE model.
IF the population mates at random, then we can use the product rule to determine
the probability of any two gametes coming together. The propability that and
'A' sperm fertilizes an 'A' egg = 0.6 x 0.6 = 0.36. And of course, this is the
only way to produce an 'AA' zygote. So, the frequency of 'Aa' zygotes (the F1
offspring) produced by this population should be 0.36. Likewise, the probability
that an 'a' sperm fertilizes an 'a' egg = 0.4 x 0.4 = 0.16. And again, this
is the only way to make an 'aa' zygote, so the total frequency of 'aa' zygotes
in the F1 will be 0.16. Now, there are two ways to make an 'Aa' zygote: an 'A'
sperm can fertilize an 'a' egg (probability = 0.6 x 0.4 = 0.24), and an 'a'
sperm can fertilize an 'A' egg (also with a probability of 0.4 x 0.6 = 0.24).
So, the total frequency of Aa zygotes in the F1 will be 2 x 0.24 = 0.48. If
we generalize, and let f(A) = p and f(a) = q, then the genotypic frequencies
under HWE can be calculated as: f(AA) = p2, f(Aa) = 2pq, and f(aa)
= q2.
Now, of course, these calculations
will only be true IF the population mates at random. AND, they will only be
true if there is no mutation. If 'A" alleles are mutating into 'a' alleles,
then the gene frequencies will not be 0.6 and 0.4, and calculations based on
these numbers will not be correct. So, we must assume NO MUTATION. Likewise,
we can't have any migration; we can't have 1000 AA individuals migrate into
our population, or that would change the gene frequencies, too; and our predictions
based on frequencies of 0.6 and 0.4 would be incorrect. So, we must assume NO
MIGRATION, too.
So, at this point we have zygotes
at the frequencies shown in the "Genotypes, F1" row. In order for
there to be no change in the genetic structure of the population, there must
be NO SELECTION. In other words, all genotypes must have the same probability
of survival and reproduction. Only then will they contribute gametes at frequencies
of p = 0.6 and q = 0.4. (If there were selection, and if AA individuals were
the only zygotes to survive to reproeuce, for instance, then the gene frequencies
would change and our predictions based on frequencies of 0.6 and 0.4 would not
be correct).
And finally, this model will only
be explicitly true for populations that are infinitely large: because that is
the only time when we can be garaunteed that predictions based on random chance
will be exactly met. (Think about it this way... suppose I give you a coin that
is absolutely perfectly balanced at the molecular level. It IS PERFECTLY BALANCED.
And suppose I ask you, "how many times do you have to flip that coin to
be ABSOLUTELY SURE of producing a 50:50 ratio of heads to tails? Well, if you
only flip it four times, you know that, just by chance, you would often get
3 heads and a tail or 3 tails and a head. And even if you flip it 10,000 times,
you might get 5001 heads and 4999 tails, even though the coin is perfectly balanced.
To be absolutely garaunteed that the predictions of this probabilisitic model
will be met exactly, you must flip the coin an infinite number of times. Obviously,
this is a theoretical constraint because no population is infinitely large.
But this is a theoretical model of no change, so we can employ theoretical expectations.
The same is true of our 'expectation' of a perfectly balanced coin - this expectation
will only be met, for sure, in an infinitely large sample. Yet we continually
employ that expectation for a perfectly balanced coin.
So, that is why these assumptions
exist. It is only when these are met that a population will not evolve. Wow.
That should seem rather amazing. It is only when these assumptions are ALL met
that a population WON'T change. If any of these assumptions is not met, a population's
genetic structure WILL change... and that is evolution. So, from this analysis,
we should expect populations to evolve - it is only under a rare combination
of events (no, mutation, no selection, no migration, random mating, and an infinitiely
large population) that evolution WON'T happen.
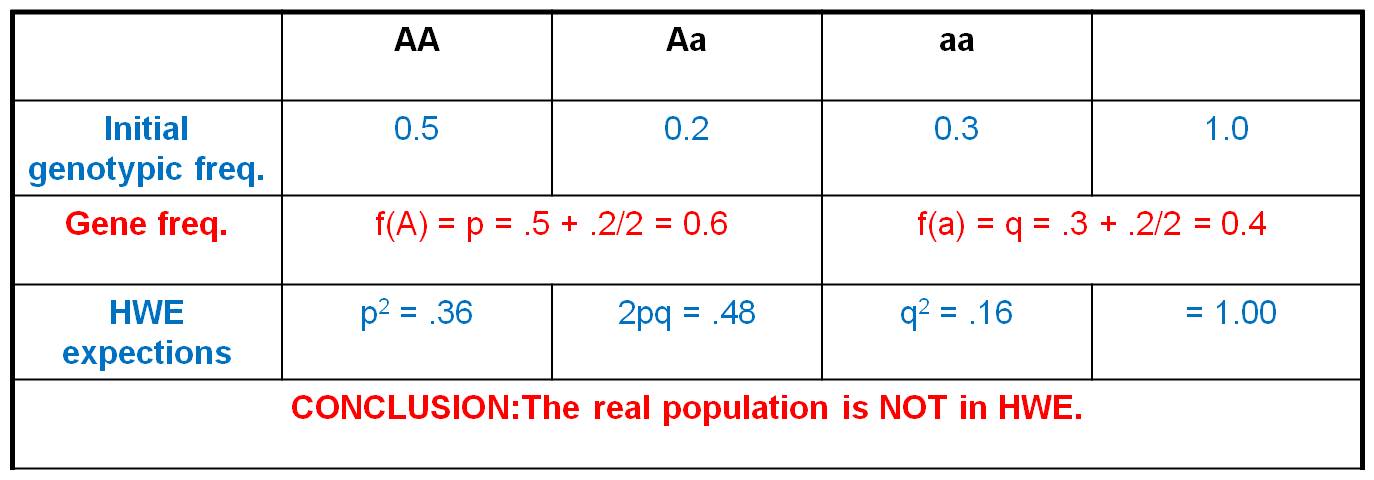
3. Utility
- If no real populations can explicitly
meet these assumptions, how can the model be useful? For instance, no
real population is infinitely large, so how can the model be useful? We
use it for COMPARISON. This model describes what the genotypic frequencies
should be IF the population was in equilibrium.
If the real genotypic frequencies are not close to these expectations, then
the population is not in HWE.... it is evolving. And if a population is not
in HWE, then the population must be violating one of the assumptions of the
HWE model. Think about that. The HWE is only 'true' if the assumptions
are being met. If your real population differs from the model, then one
of the assumptions must not apply to your real population. This narrows
your focus on WHY the real populations isn't behaving randomly... and it might
identify WHY the population is evolving.... which is a biologically interesting
question.
 -
Again, the coin analogy applies. No REAL coin is probably exactly perfectly
balanced. But, if I give you a coin and ask you how balanced it is, you flip
it a few times and compare its behavior to WHAT YOU WOULD EXPECT FROM A PERFECT
COIN (50:50 RATIO). Even though a perfectly balanced coin does not exist, we
can use this theoretical model as a benchmark, to compare the behavior of real
coins. Many real coins act in a manner that is consistent enough with
the expectations from a perfectly balanced coin that we are willing to use them
AS IF they were perfectly balanced. The Hardy Weinberg Equilibrium
Model is the same... it is a model of no change against which we can measure
real populations.
-
Again, the coin analogy applies. No REAL coin is probably exactly perfectly
balanced. But, if I give you a coin and ask you how balanced it is, you flip
it a few times and compare its behavior to WHAT YOU WOULD EXPECT FROM A PERFECT
COIN (50:50 RATIO). Even though a perfectly balanced coin does not exist, we
can use this theoretical model as a benchmark, to compare the behavior of real
coins. Many real coins act in a manner that is consistent enough with
the expectations from a perfectly balanced coin that we are willing to use them
AS IF they were perfectly balanced. The Hardy Weinberg Equilibrium
Model is the same... it is a model of no change against which we can measure
real populations.
If HWE can be assumed, then the frequency
of recessive diseases can be assumed to equal q2, and the frequency of carriers
in the population can be estimated like this:
1) The frequency of hemachromatosis
worldwide is 1/450. If we assume that hemochromatosis is caused by a recessive
gene (q), and if we assume the population is in HWE with respect to this trait,
then q2 = 1/450 = 0.002. So, we take the square-root of both sides
to find q = 0.047. Well, if q = 0.047, and if p + q = 1, then p = 1 - 0.047
= 0.953.
2) If q = 0.047 and p = 0.953, then
the frequency of heterozygous carriers = 2pq = 0.09. So, we estimate that 9%
of the population are carriers.
Now, you might say, "but we
just determined that HWE would be unusual; so why would we assume it is
true for a given gene?" Well, a deleterious gene has already been
largely weeded out of a population, so selection against the few alleles that
are left is really weak. Indeed, this condition may not influence reproductive
success, anyway (NO SELECTION). In addition, we don't select mates based on
whether they have hemochromatosis (I bet you NEVER asked your date if they have
hemochromatosis, for example!!), so we can assume there is RANDOM MATING in
the population with respect to this trait. And although the human population
is not infinite, it is really big (6.9 BILLION), so the effect of sampling error
is probably very small. Mutation is very rare, so the effects of mutation are
likely to be very small. And if we are making an estimate based on the whole
human population, then there can be no 'migrants' coming in from somewhere else
(Martians?). So, in some cases, we can reasonably assume a population might
be in HWE for a given gene. Of course, we could be wrong... and we would test
that prediction by sampling individuals in the population and determining the
frequency of heterozygotes genetically. But at least we would have a working
hypothesis.
Study
Questions:
1. What
is Platonic essentialism (or 'idealism') and how did it hinder the consideration
of evolutionary ideas?
2. What contributions
did Copernicus, Kepler, Galileo, and Newton make to our understanding of the
solar system? Frame these in the context of the scientific method.
3. What counter-intuitive
effect did the renaissance have on the development of modern (evolutionary)
biology?
4. What is the
principle of Natural Theology, as professed by Aquinas and Paley?
5. Why did
Lamarck conclude that species change over time, and what philosophical dilemma
was he trying to solve?
6. What is "uniformitarianism" and how was it important to the development of Darwin's
ideas?
7. What observations
did Hutton make, and what did he conclude from these observations?
8. What two patterns
occur in the fossil record that impress Darwin regarding the hypothesis of evolution
and common descent?
9. What are homologous
structures? What correlations occurs with the environment?
10. What are analogous
structures? What correlation occurs with the environment?
11. What are
vestigial structures, and why were they so important to Darwin's refutation
of Paley?
12. How did
Darwin explain the existence of 'convergent communities"?
13. The Galapagos
are dominated by many unique species of reptiles, birds, and marine mammals. What did this
non-random assemblage suggest to Darwin about their origin, and how was evolution
implied?
14. Why were
the mockingbirds so critical to Darwin's ideas about the production of new species?
15. How did Darwin
use the comparative method and observations of 'artificial selection' to produce
the theory of 'natural selection'?
16. How were Malthus's
observations and conclusion rlevant to the development of Darwin's theory?
17. Outline the
theory of natural selection as an argument, with three premises, 3 conclusions,
and a corollary.
18.
What are the five assumptions of the Hardy-Weinberg Equilibrium Model?
19.
Consider the following population:
| |
AA |
Aa |
aa |
| Number
of Individuals |
60 |
20 |
20 |
- -
calculate the genotypic frequencies.
- -
calculate the gene frequencies
- -
calculate the HARDY WEINBERG EQUILIBRIUM frequencies.
20.
If the HWE model does not describe any real population, how can it be useful?
 B. Darwin's Contributions: The
Origin of Species (1859)
B. Darwin's Contributions: The
Origin of Species (1859)